

Naringenin Inhibition of the Pseudomonas aeruginosa Quorum Sensing Response Is Based on Its Time-Dependent Competition With N-(3-Oxo-dodecanoyl)-L-homoserine Lactone for LasR Binding
- PMID: 32181260
- PMCID: PMC7059128
- DOI: 10.3389/fmolb.2020.00025
Naringenin Inhibition of the Pseudomonas aeruginosa Quorum Sensing Response Is Based on Its Time-Dependent Competition With N-(3-Oxo-dodecanoyl)-L-homoserine Lactone for LasR Binding
Abstract
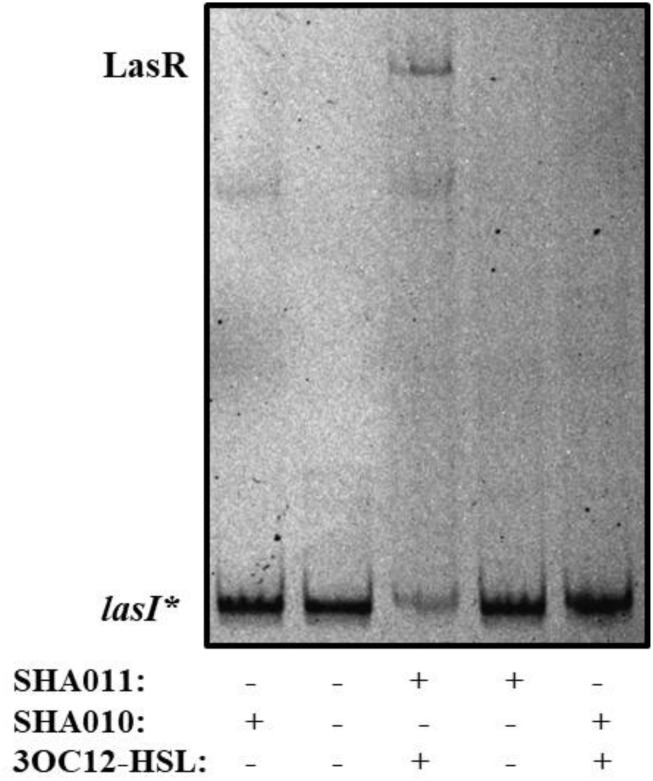
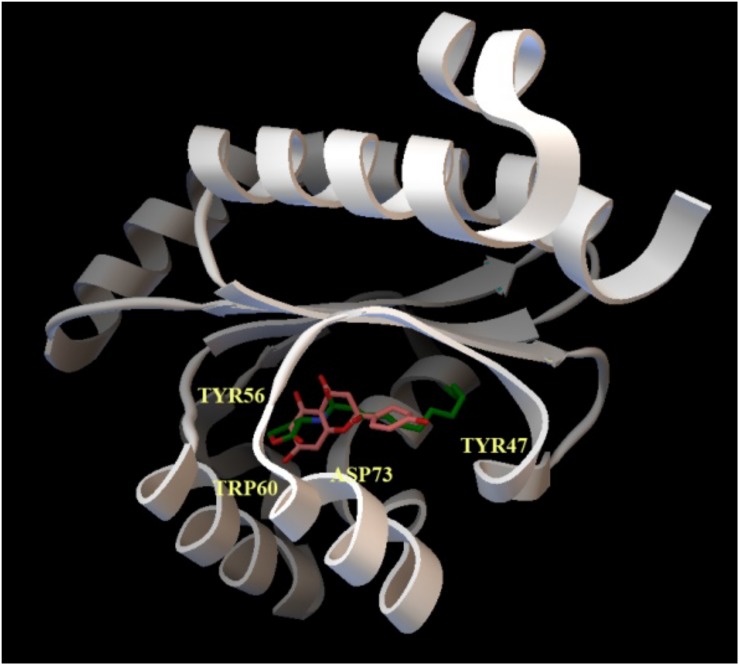
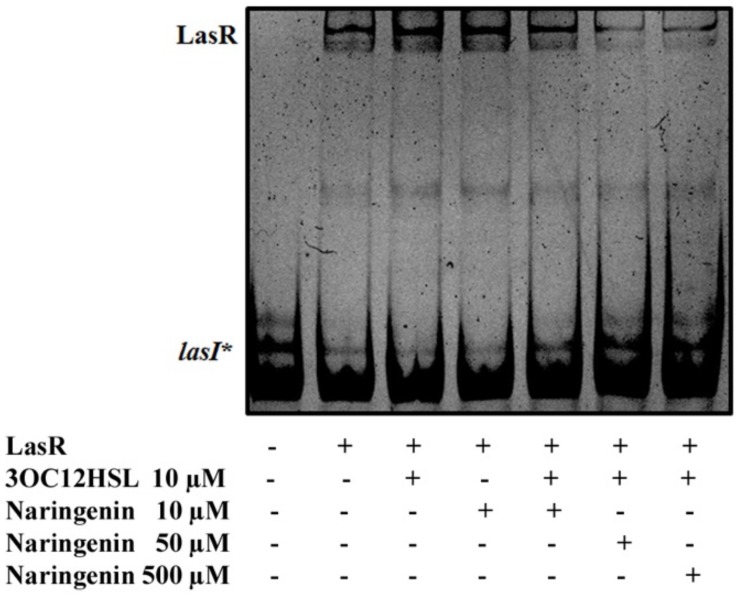
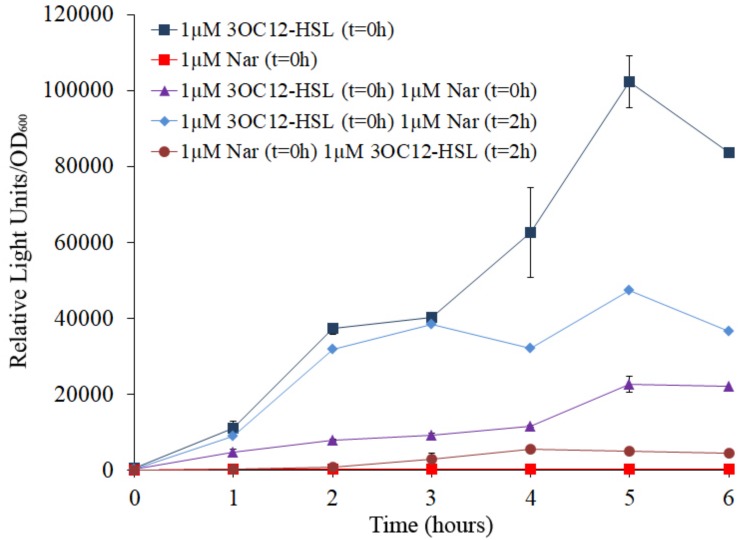
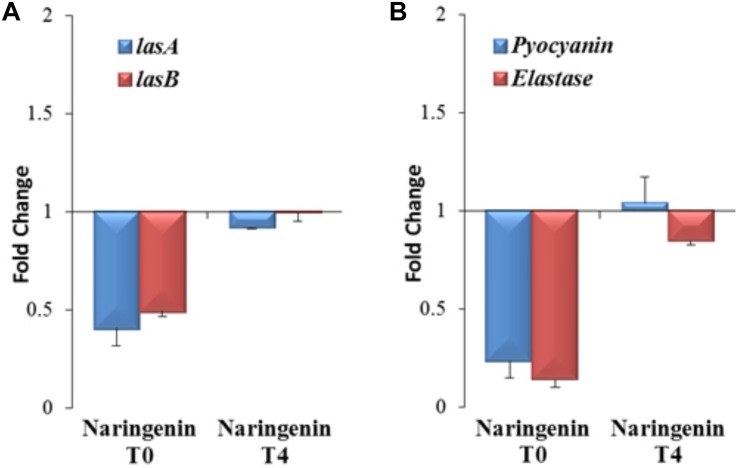
Bacterial quorum sensing (QS) is a cell-to-cell communication system that governs the expression of a large set of genes involved in bacterial-host interactions, including the production of virulence factors. Conversely, the hosts can produce anti-QS compounds to impair virulence of bacterial pathogens. One of these inhibitors is the plant flavonoid naringenin, which impairs the production of QS-regulated Pseudomonas aeruginosa virulence factors. In the present work, we analyze the molecular basis for such inhibition. Our data indicate that naringenin produces its effect by directly binding the QS regulator LasR, hence competing with its physiological activator, N-(3-oxo-dodecanoyl)-L-homoserine lactone (3OC12-HSL). The in vitro analysis of LasR binding to its cognate target DNA showed that the capacity of naringenin to outcompete 3OC12-HSL, when the latter is previously bound to LasR, is low. By using an E. coli LasR-based biosensor strain, which does not produce 3OC12-HSL, we determined that the inhibition of LasR is more efficient when naringenin binds to nascent LasR than when this regulator is already activated through 3OC12-HSL binding. According to these findings, at early exponential growth phase, when the amount of 3OC12-HSL is low, naringenin should proficiently inhibit the P. aeruginosa QS response, whereas at later stages of growth, once 3OC12-HSL concentration reaches a threshold enough for binding LasR, naringenin would not efficiently inhibit the QS response. To test this hypothesis, we analyze the potential effect of naringenin over the QS response by adding naringenin to P. aeruginosa cultures at either time zero (early inhibition) or at stationary growth phase (late inhibition). In early inhibitory conditions, naringenin inhibited the expression of QS-regulated genes, as well as the production of the QS-regulated virulence factors, pyocyanin and elastase. Nevertheless, in late inhibitory conditions, the P. aeruginosa QS response was not inhibited by naringenin. Therefore, this time-dependent inhibition may compromise the efficiency of this flavonoid, which will be effective just when used against bacterial populations presenting low cellular densities, and highlight the importance of searching for QS inhibitors whose mechanism of action does not depend on the QS status of the population.
Keywords: LasR; Pseudomonas aeruginosa; naringenin; quorum sensing; virulence inhibitor.
Copyright © 2020 Hernando-Amado, Alcalde-Rico, Gil-Gil, Valverde and Martínez.
Figures
References
-
- Bacha K., Tariku Y., Gebreyesus F., Zerihun S., Mohammed A., Weiland-Brauer N., et al. (2016). Antimicrobial and anti-Quorum sensing activities of selected medicinal plants of Ethiopia: implication for development of potent antimicrobial agents. BMC Microbiol. 16:139. 10.1186/s12866-016-0765-9 - DOI - PMC - PubMed
-
- Chatterjee A., Cui Y., Yang H., Collmer A., Alfano J. R., Chatterjee A. K. (2003). GacA, the response regulator of a two-component system, acts as a master regulator in Pseudomonas syringae pv. tomato DC3000 by controlling regulatory RNA, transcriptional activators, and alternate sigma factors. Mol. Plant Microbe Interact. 16 1106–1117. 10.1094/mpmi.2003.16.12.1106 - DOI - PubMed