

Emerging roles of novel small non-coding regulatory RNAs in immunity and cancer
- PMID: 32186461
- PMCID: PMC7549716
- DOI: 10.1080/15476286.2020.1737442
Emerging roles of novel small non-coding regulatory RNAs in immunity and cancer
Abstract
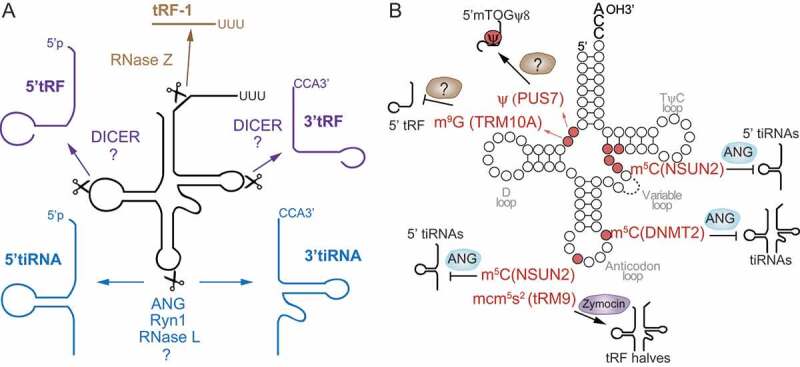
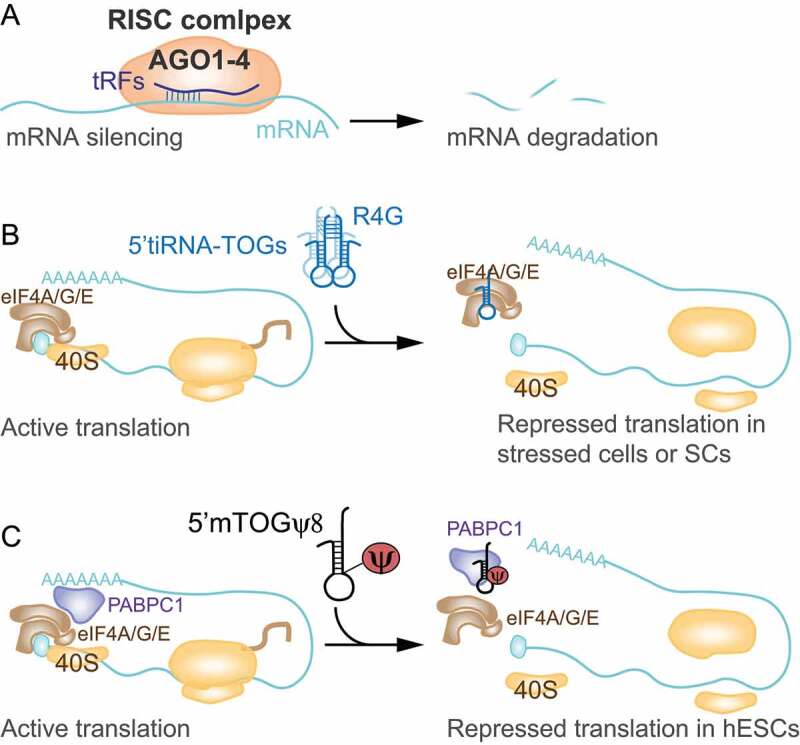
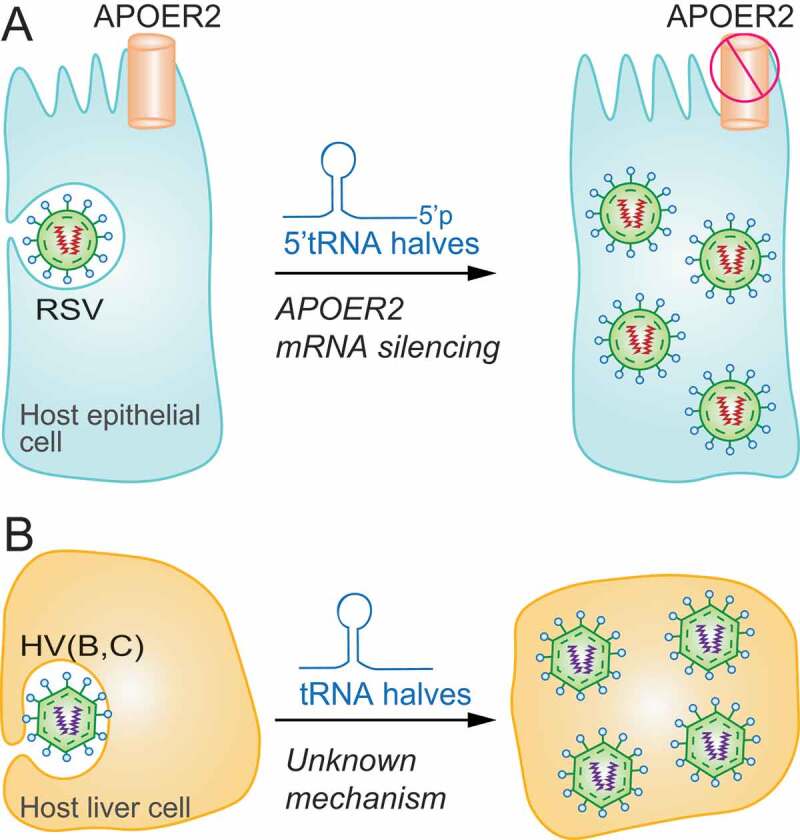
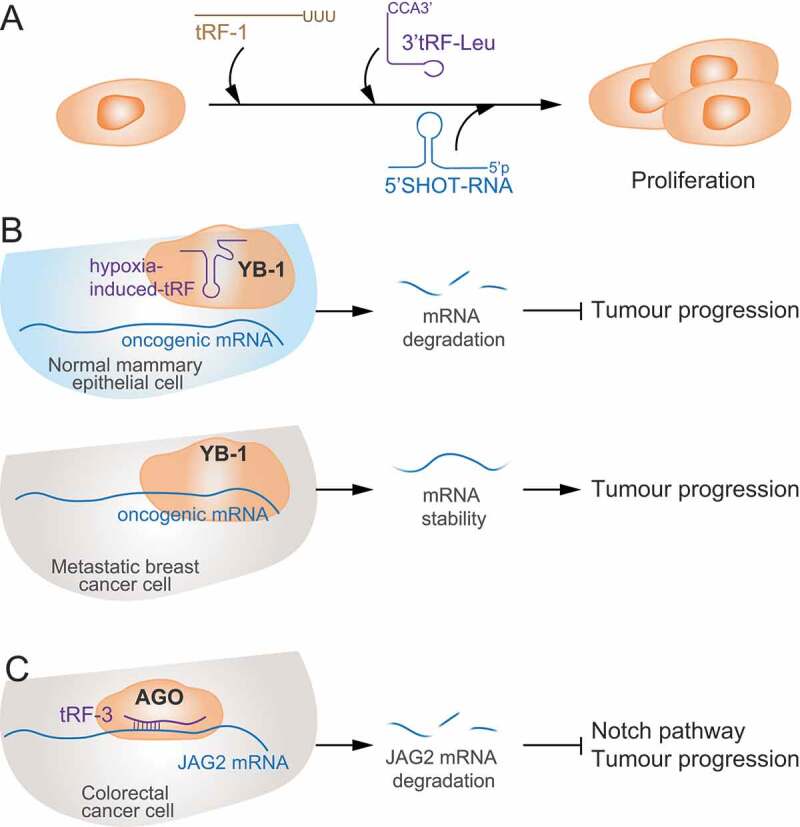
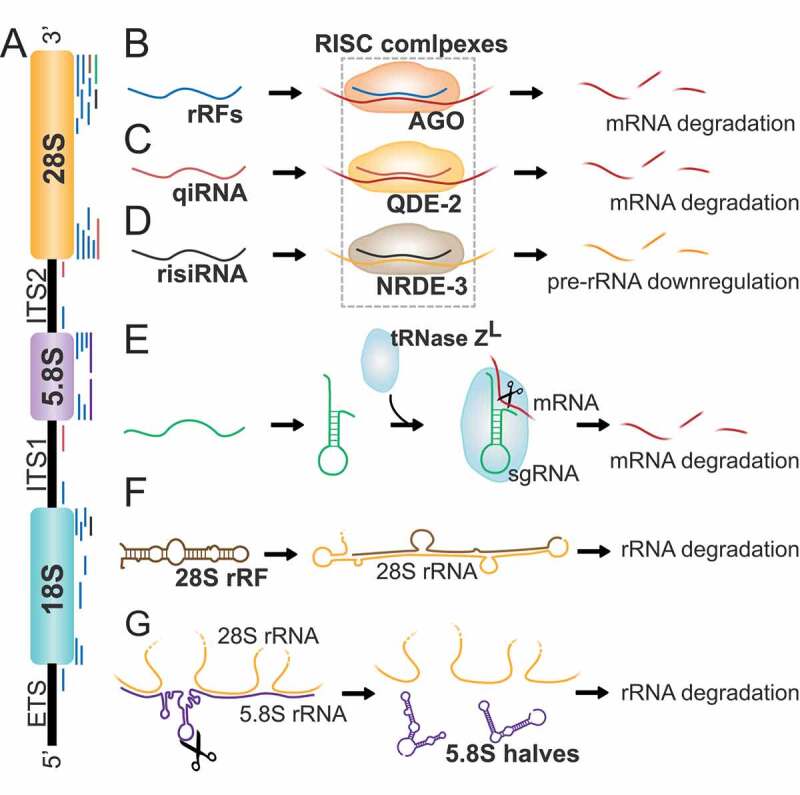
The term small non-coding RNAs (ncRNAs) refers to all those RNAs that even without encoding for a protein, can play important functional roles. Transfer RNA and ribosomal RNA-derived fragments (tRFs and rRFs, respectively) are an emerging class of ncRNAs originally considered as simple degradation products, which though play important roles in stress responses, signalling, or gene expression. They control all levels of gene expression regulating transcription and translation and affecting RNA processing and maturation. They have been linked to pivotal cellular processes such as self-renewal, differentiation, and proliferation. For this reason, mis-regulation of this novel class of ncRNAs can lead to various pathological processes such as neurodegenerative and development diseases, metabolism and immune system disorders, and cancer. In this review, we summarise the classification, biogenesis, and functions of tRFs and rRFs with a special focus on their role in immunity and cancer.
Keywords: Transfer RNA fragments; cancer; immune system; rRFs; ribosomal RNA fragments; stress-induced tRNA cleavage; tRFs; tRNA cleavage; translation initiation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Mattick JS, Makunin IV.. Non-coding RNA. Hum Mol Genet. 2006;15(Spec No 1):R17–29. - PubMed
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854. - PubMed
-
- Cech TR, Steitz JA. The noncoding RNA revolution-trashing old rules to forge new ones. Cell. 2014;157:77–94. - PubMed
-
- Borek E, Baliga BS, Gehrke CW, et al. High turnover rate of transfer RNA in tumor tissue. Cancer Res. 1977;37:3362–3366. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical