

Diapedesis-Induced Integrin Signaling via LFA-1 Facilitates Tissue Immunity by Inducing Intrinsic Complement C3 Expression in Immune Cells
- PMID: 32187519
- PMCID: PMC7111494
- DOI: 10.1016/j.immuni.2020.02.006
Diapedesis-Induced Integrin Signaling via LFA-1 Facilitates Tissue Immunity by Inducing Intrinsic Complement C3 Expression in Immune Cells
Abstract
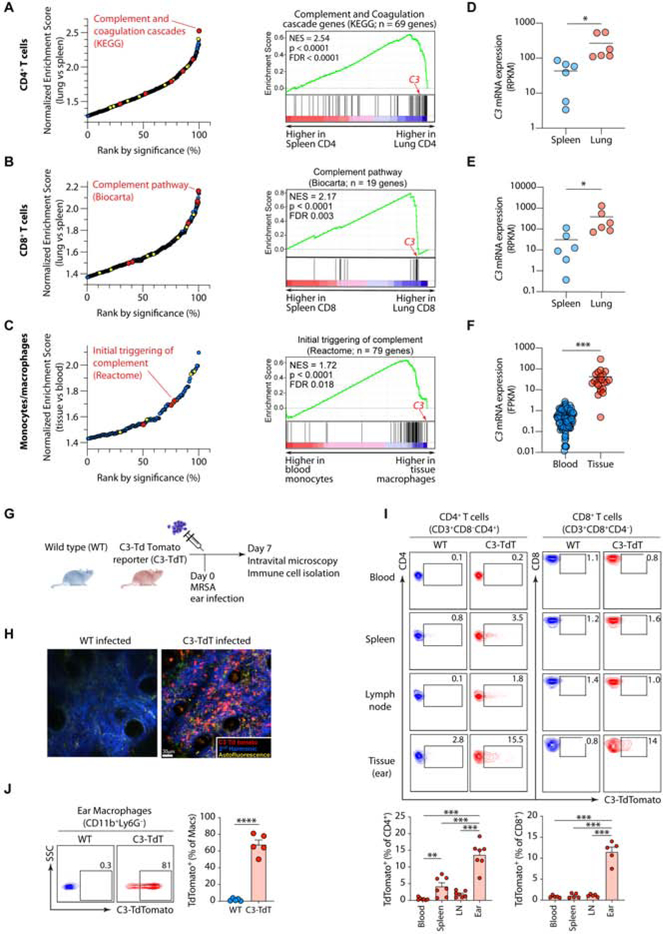
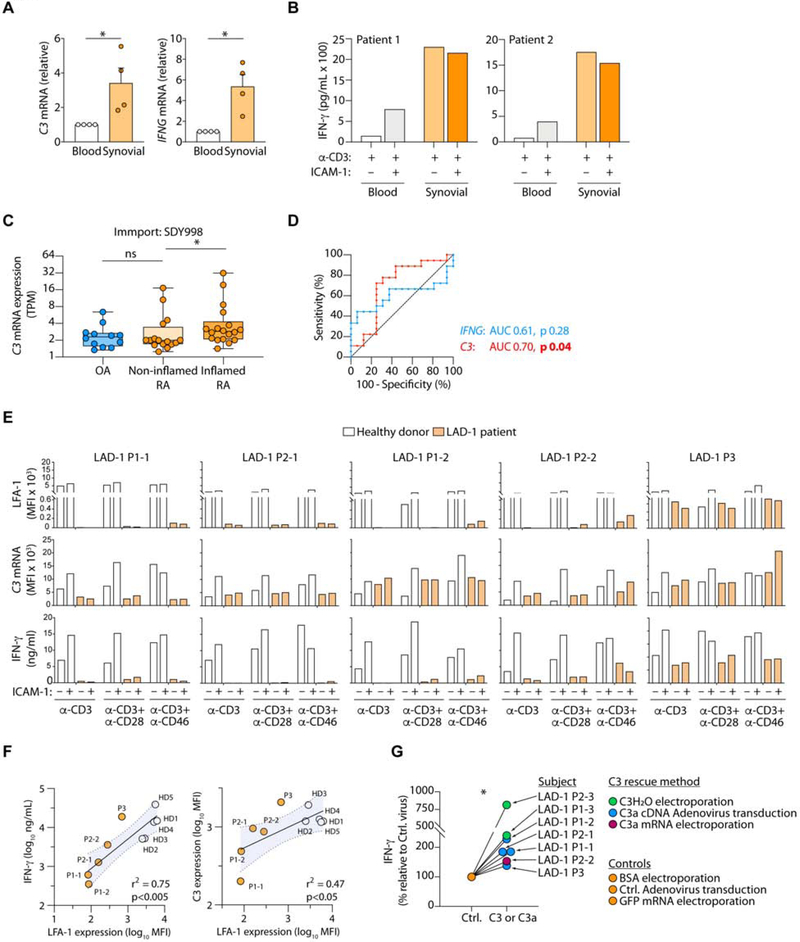
Intrinsic complement C3 activity is integral to human T helper type 1 (Th1) and cytotoxic T cell responses. Increased or decreased intracellular C3 results in autoimmunity and infections, respectively. The mechanisms regulating intracellular C3 expression remain undefined. We identified complement, including C3, as among the most significantly enriched biological pathway in tissue-occupying cells. We generated C3-reporter mice and confirmed that C3 expression was a defining feature of tissue-immune cells, including T cells and monocytes, occurred during transendothelial diapedesis, and depended on integrin lymphocyte-function-associated antigen 1 (LFA-1) signals. Immune cells from patients with leukocyte adhesion deficiency type 1 (LAD-1) had reduced C3 transcripts and diminished effector activities, which could be rescued proportionally by intracellular C3 provision. Conversely, increased C3 expression by T cells from arthritis patients correlated with disease severity. Our study defines integrins as key controllers of intracellular complement, demonstrates that perturbations in the LFA-1-C3-axis contribute to primary immunodeficiency, and identifies intracellular C3 as biomarker of severity in autoimmunity.
Keywords: LAD-1; LFA-1; T cells; Th1 cells; autoimmunity; biomarker; complosome; integrins; intracellular complement; lymphocyte adhesion deficiency type 1; metabolism.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures




References
-
- Abraham C, Griffith J, and Miller J (1999). The dependence for leukocyte function-associated antigen-1/ICAM-1 interactions in T cell activation cannot be overcome by expression of high density TCR ligand. J Immunol 162, 4399–4405. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- ZIA HL006222/ImNIH/Intramural NIH HHS/United States
- 097261/Z/11/Z/WT_/Wellcome Trust/United Kingdom
- K22 HL125593/HL/NHLBI NIH HHS/United States
- 102932/Z/13/Z/WT_/Wellcome Trust/United Kingdom
- 21226/VAC_/Versus Arthritis/United Kingdom
- R44 DK075149/DK/NIDDK NIH HHS/United States
- R43 DK075149/DK/NIDDK NIH HHS/United States
- MR/M004600/1/MRC_/Medical Research Council/United Kingdom
- ZIA DK075149/ImNIH/Intramural NIH HHS/United States
- DH_/Department of Health/United Kingdom
- MR/J006742/1/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- ZIA HL006223/ImNIH/Intramural NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
