

A metabolic switch regulates the transition between growth and diapause in C. elegans
- PMID: 32188449
- PMCID: PMC7081555
- DOI: 10.1186/s12915-020-0760-3
A metabolic switch regulates the transition between growth and diapause in C. elegans
Abstract
Background: Metabolic activity alternates between high and low states during different stages of an organism's life cycle. During the transition from growth to quiescence, a major metabolic shift often occurs from oxidative phosphorylation to glycolysis and gluconeogenesis. We use the entry of Caenorhabditis elegans into the dauer larval stage, a developmentally arrested stage formed in response to harsh environmental conditions, as a model to study the global metabolic changes and underlying molecular mechanisms associated with growth to quiescence transition.
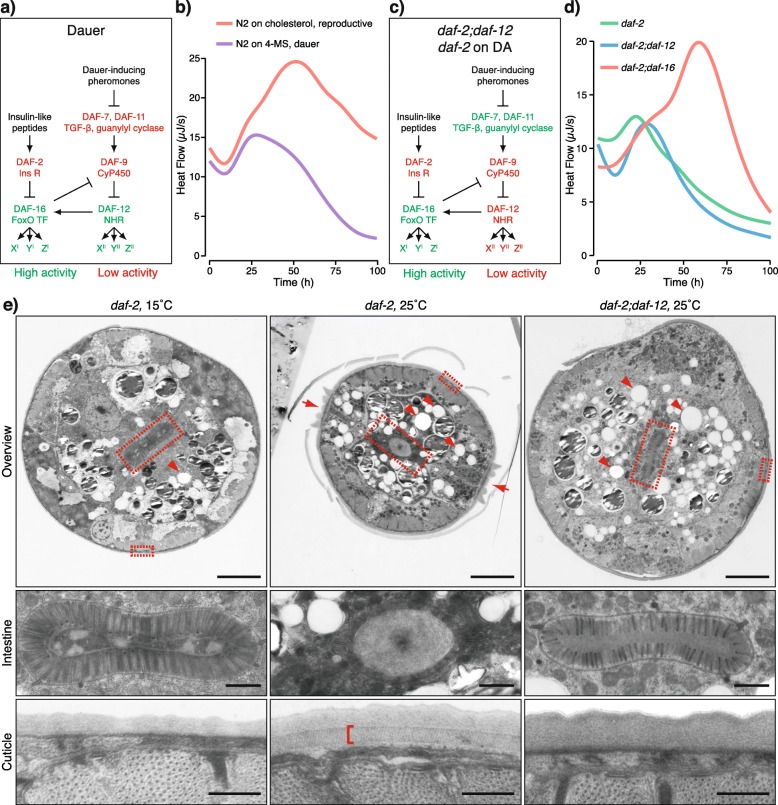
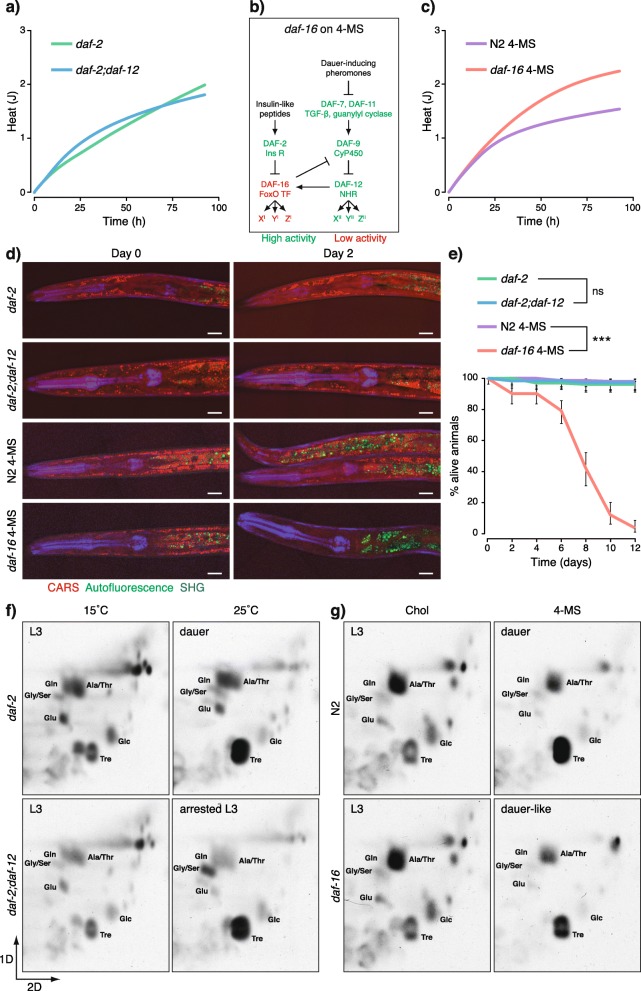
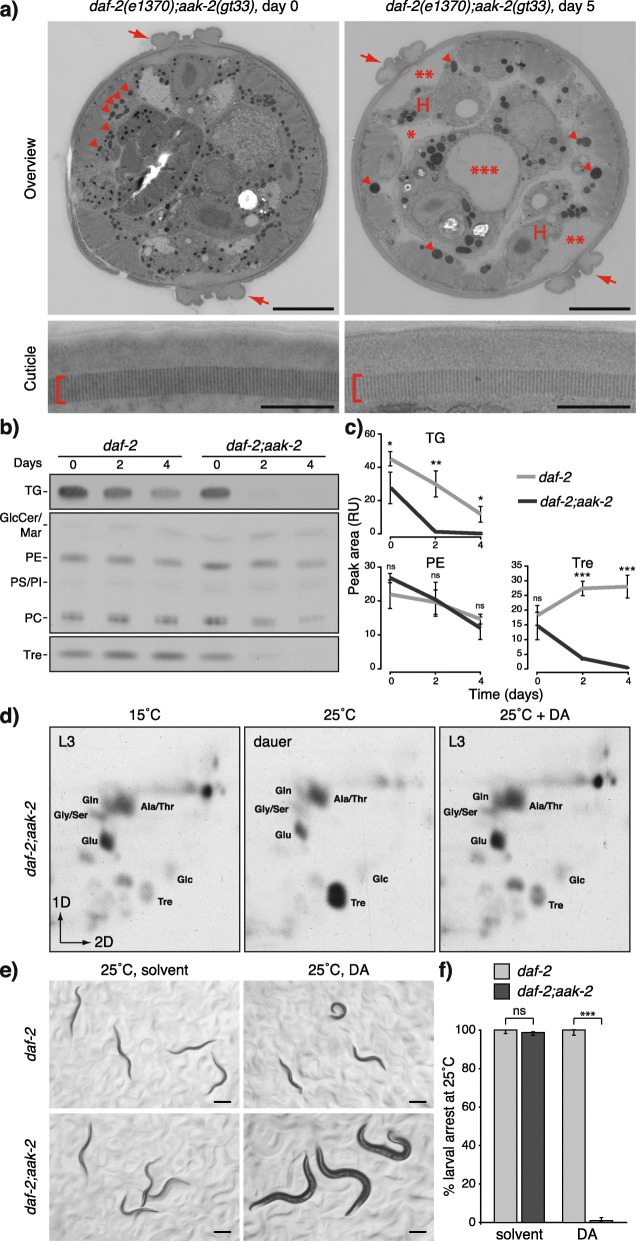
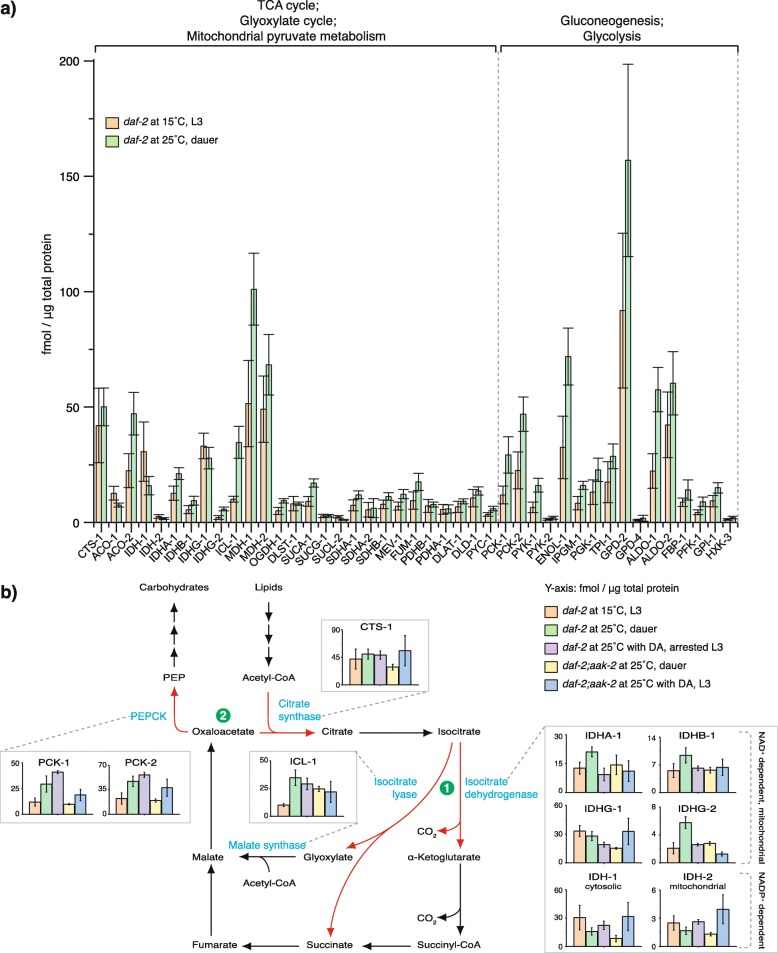
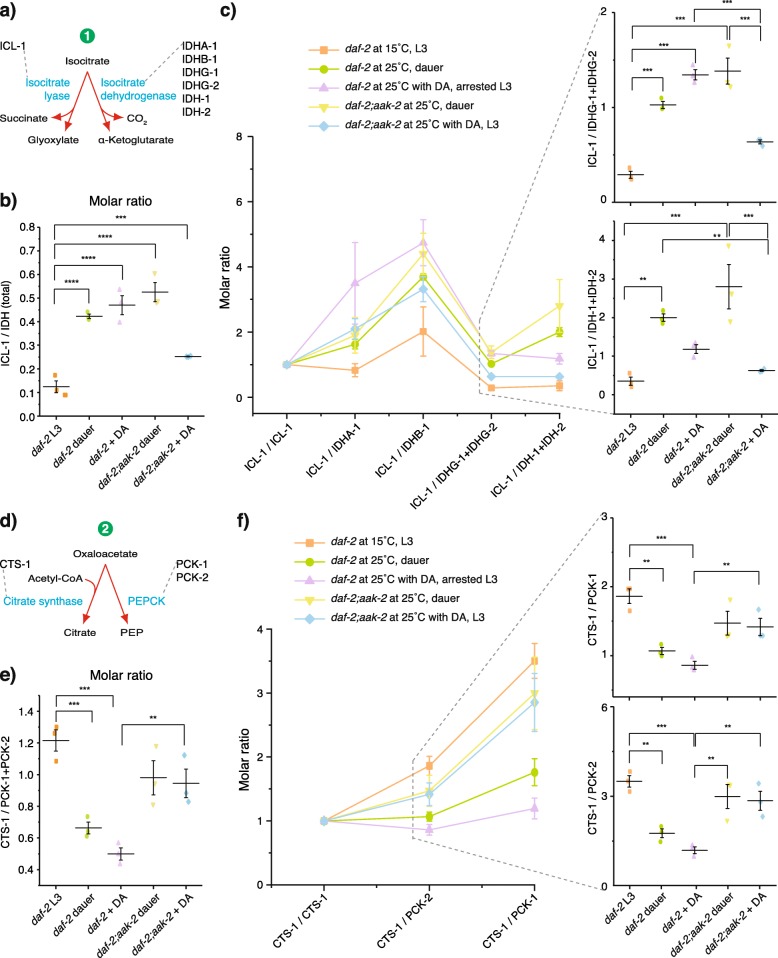
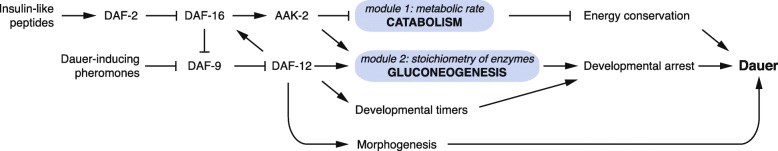
Results: Here, we show that the metabolic switch involves the concerted activity of several regulatory pathways. Whereas the steroid hormone receptor DAF-12 controls dauer morphogenesis, the insulin pathway maintains low energy expenditure through DAF-16/FoxO, which also requires AAK-2/AMPKα. DAF-12 and AAK-2 separately promote a shift in the molar ratios between competing enzymes at two key branch points within the central carbon metabolic pathway diverting carbon atoms from the TCA cycle and directing them to gluconeogenesis. When both AAK-2 and DAF-12 are suppressed, the TCA cycle is active and the developmental arrest is bypassed.
Conclusions: The metabolic status of each developmental stage is defined by stoichiometric ratios within the constellation of metabolic enzymes driving metabolic flux and controls the transition between growth and quiescence.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Smith DG, Sturmey RG. Parallels between embryo and cancer cell metabolism. Biochem Soc Trans. 2013;41(2):664–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
