

SUMOylation in development and neurodegeneration
- PMID: 32188601
- PMCID: PMC7097199
- DOI: 10.1242/dev.175703
SUMOylation in development and neurodegeneration
Abstract
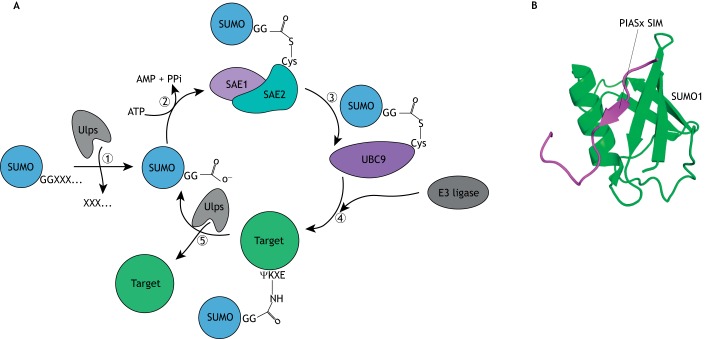
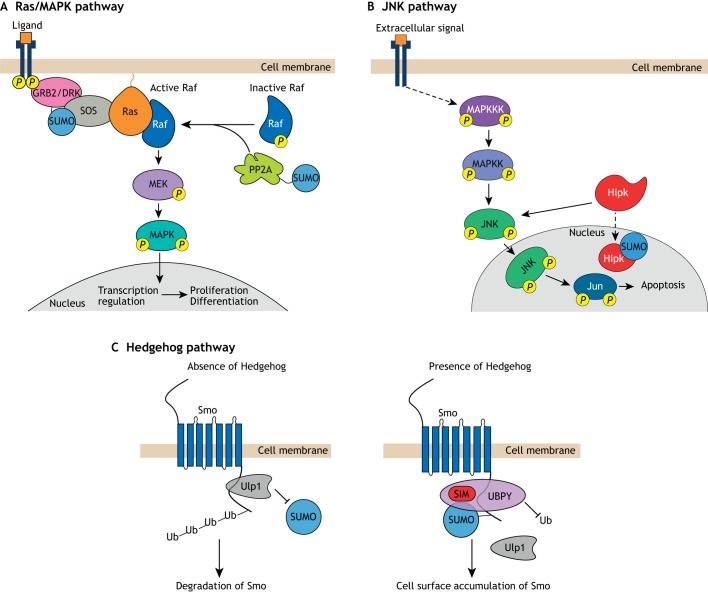
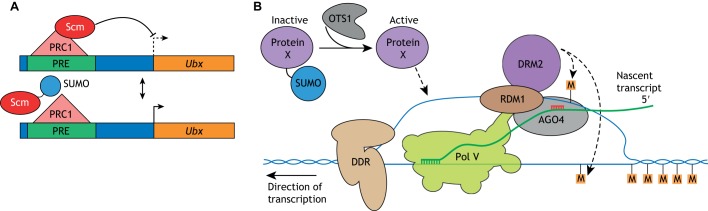
In essentially all eukaryotes, proteins can be modified by the attachment of small ubiquitin-related modifier (SUMO) proteins to lysine side chains to produce branched proteins. This process of 'SUMOylation' plays essential roles in plant and animal development by altering protein function in spatially and temporally controlled ways. In this Primer, we explain the process of SUMOylation and summarize how SUMOylation regulates a number of signal transduction pathways. Next, we discuss multiple roles of SUMOylation in the epigenetic control of transcription. In addition, we evaluate the role of SUMOylation in the etiology of neurodegenerative disorders, focusing on Parkinson's disease and cerebral ischemia. Finally, we discuss the possibility that SUMOylation may stimulate survival and neurogenesis of neuronal stem cells.
Keywords: Epigenetics; Neurodegenerative disorder; Post-translational protein modification; SUMO; Signal transduction; Ubiquitin-like protein.
© 2020. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
