

Tropism, pathology, and transmission of equine parvovirus-hepatitis
- PMID: 32192415
- PMCID: PMC7144241
- DOI: 10.1080/22221751.2020.1741326
Tropism, pathology, and transmission of equine parvovirus-hepatitis
Abstract
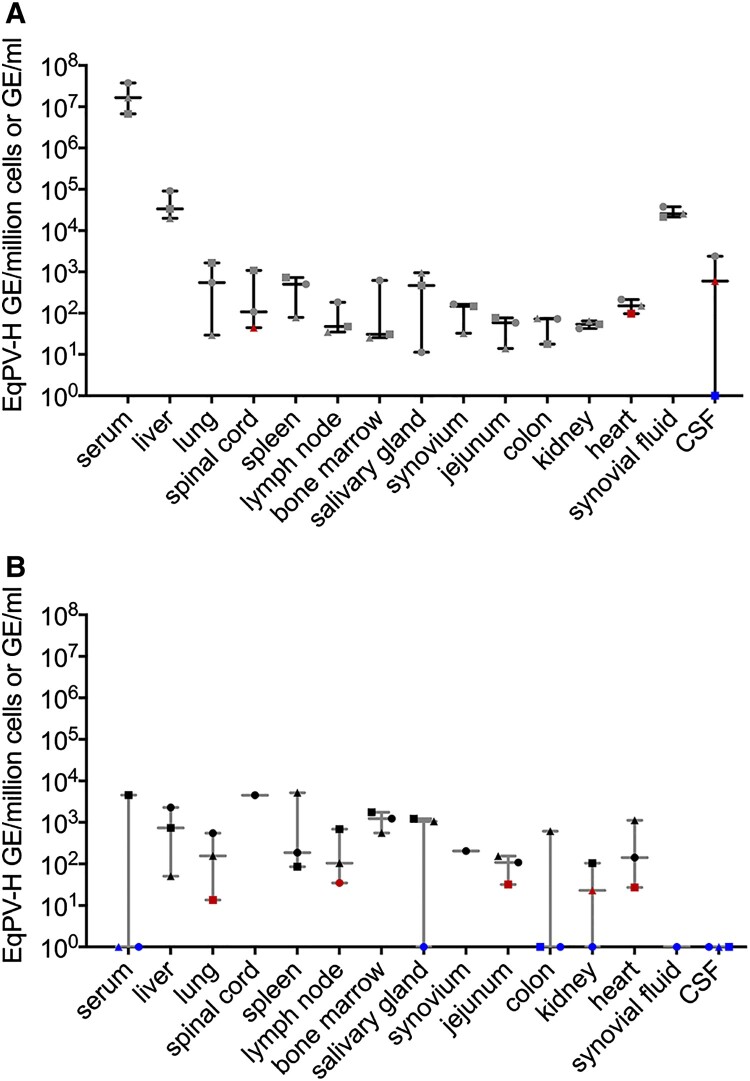
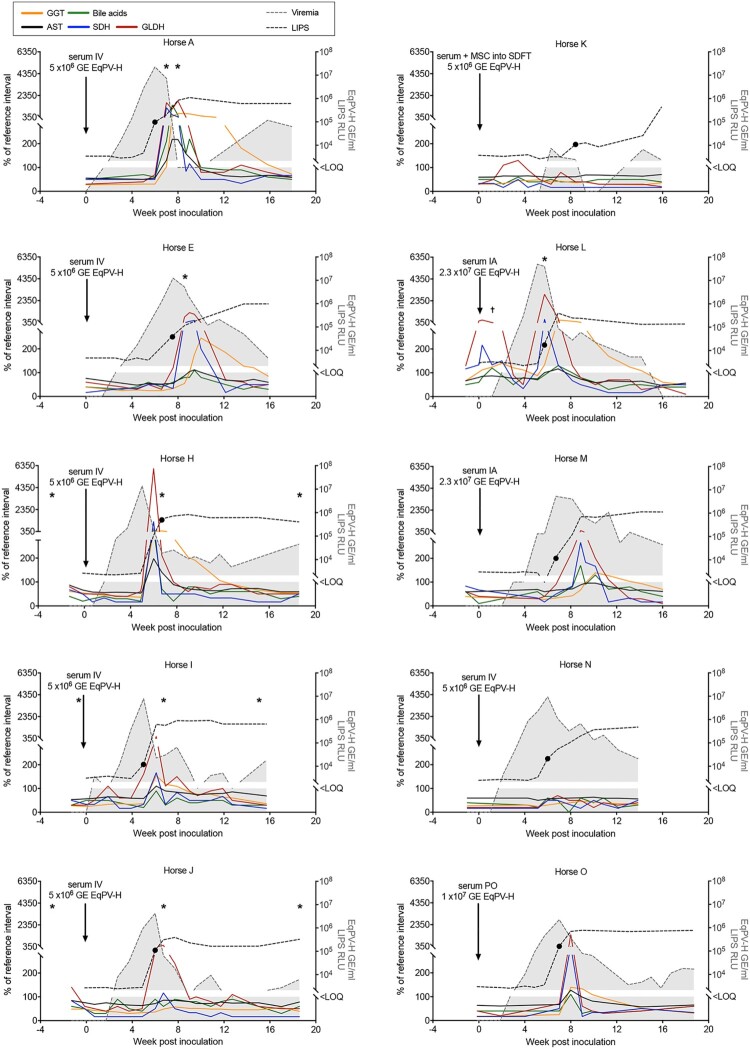
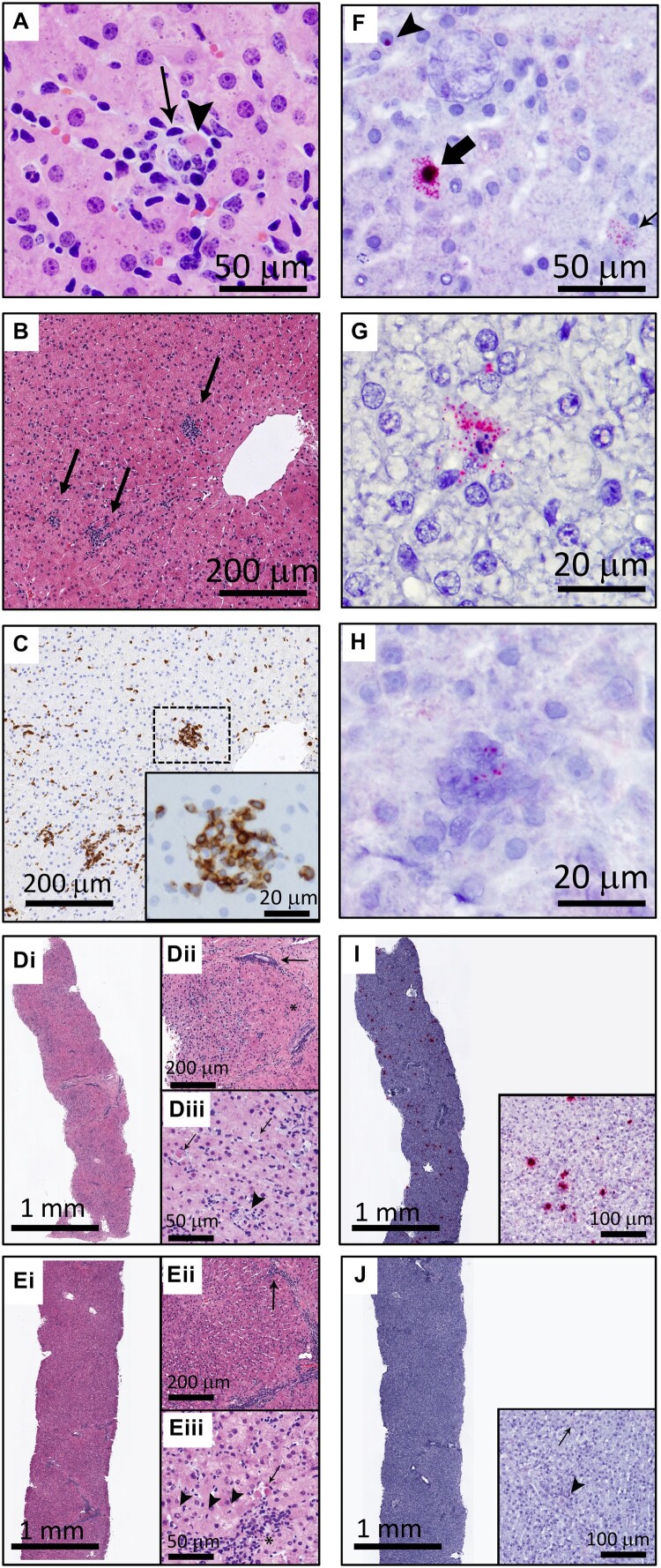
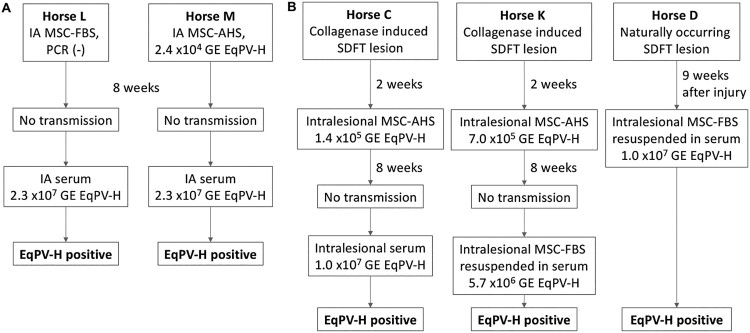
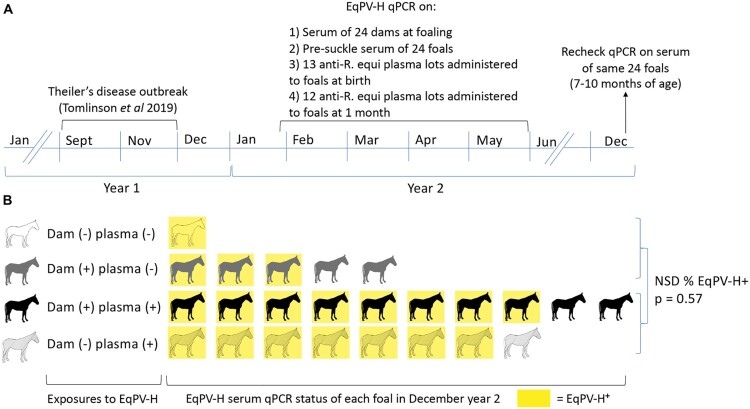
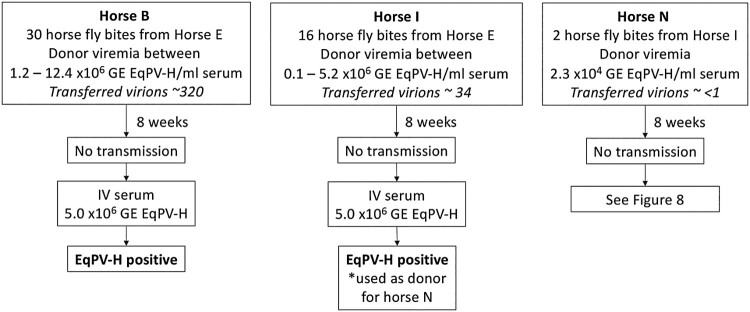
Equine parvovirus-hepatitis (EqPV-H) has recently been associated with cases of Theiler's disease, a form of fulminant hepatic necrosis in horses. To assess whether EqPV-H is the cause of Theiler's disease, we first demonstrated hepatotropism by PCR on tissues from acutely infected horses. We then experimentally inoculated horses with EqPV-H and 8 of 10 horses developed hepatitis. One horse showed clinical signs of liver failure. The onset of hepatitis was temporally associated with seroconversion and a decline in viremia. Liver histology and in situ hybridization showed lymphocytic infiltrates and necrotic EqPV-H-infected hepatocytes. We next investigated potential modes of transmission. Iatrogenic transmission via allogeneic stem cell therapy for orthopedic injuries was previously suggested in a case series of Theiler's disease, and was demonstrated here for the first time. Vertical transmission and mechanical vectoring by horse fly bites could not be demonstrated in this study, potentially due to limited sample size. We found EqPV-H shedding in oral and nasal secretions, and in feces. Importantly, we could demonstrate EqPV-H transmission via oral inoculation with viremic serum. Together, our findings provide additional information that EqPV-H is the likely cause of Theiler's disease and that transmission of EqPV-H occurs via both iatrogenic and natural routes.
Keywords: Theiler’s disease; horse fly; serum hepatitis; stem cells; vertical transmission.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures


References
-
- Lu G, Sun L, Ou J, et al. Identification and genetic characterization of a novel parvovirus associated with serum hepatitis in horses in China. Emerg Microbes Infect [Internet]. 2018;7:170, Available from: http://www.nature.com/articles/s41426-018-0174-2. - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources