

Centromere scission drives chromosome shuffling and reproductive isolation
- PMID: 32193338
- PMCID: PMC7149388
- DOI: 10.1073/pnas.1918659117
Centromere scission drives chromosome shuffling and reproductive isolation
Abstract
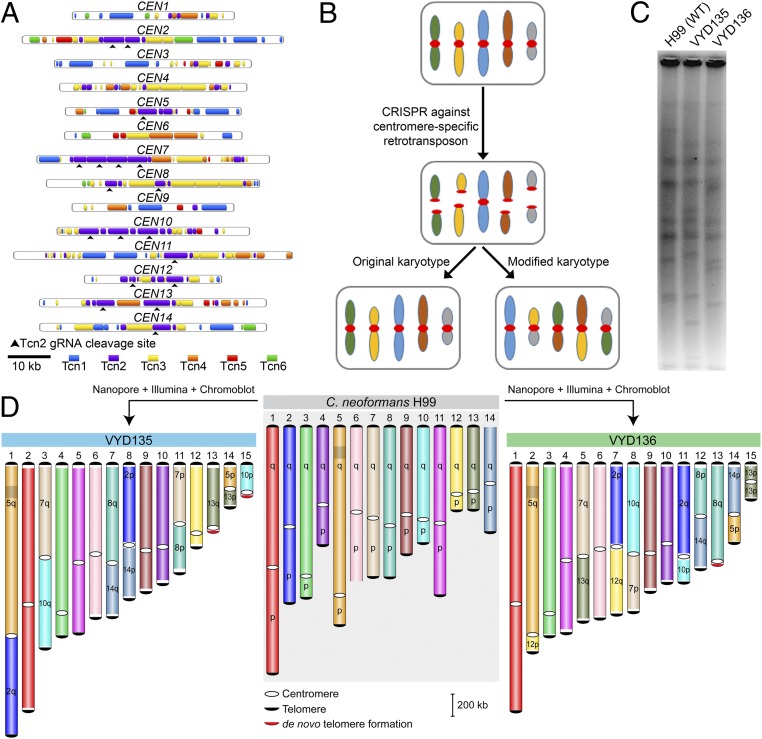
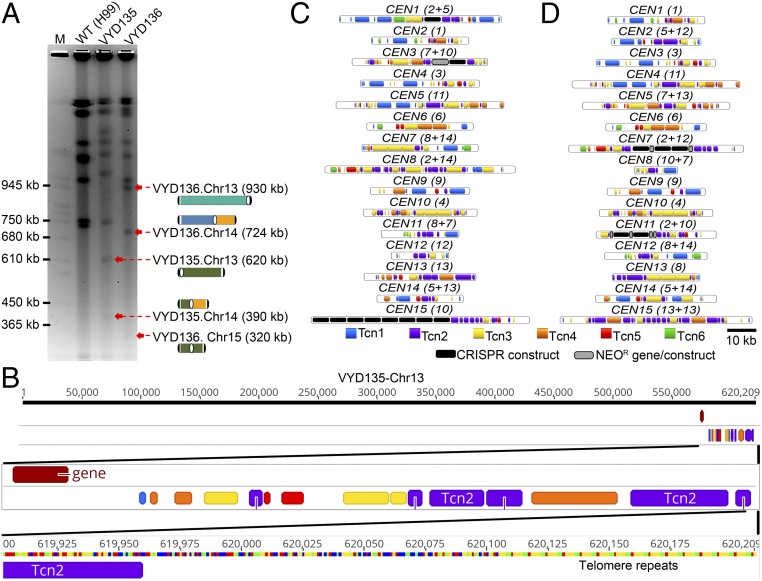
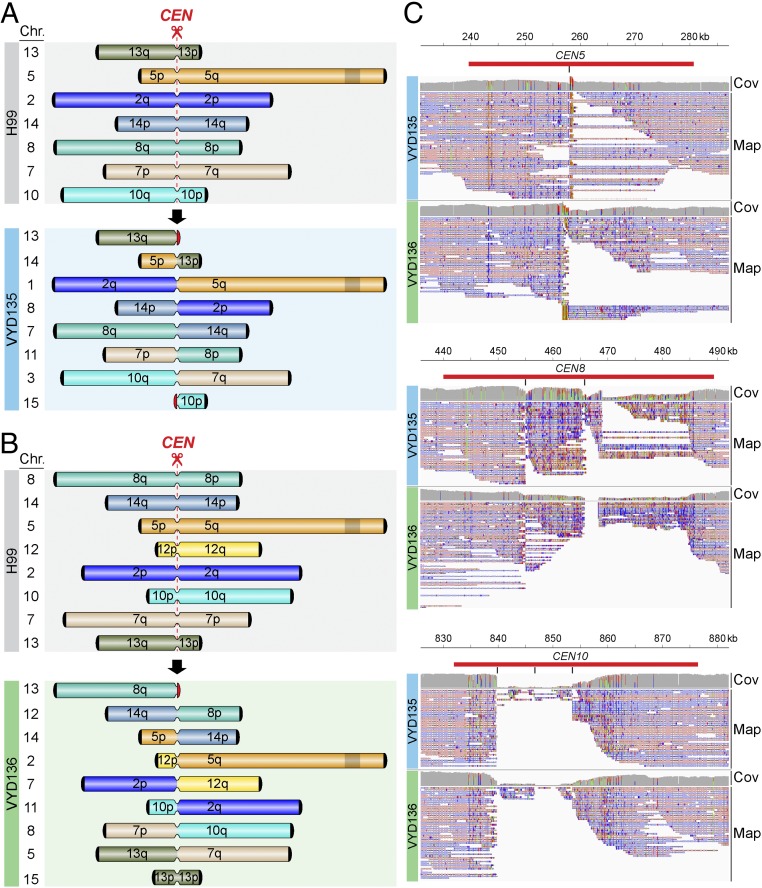
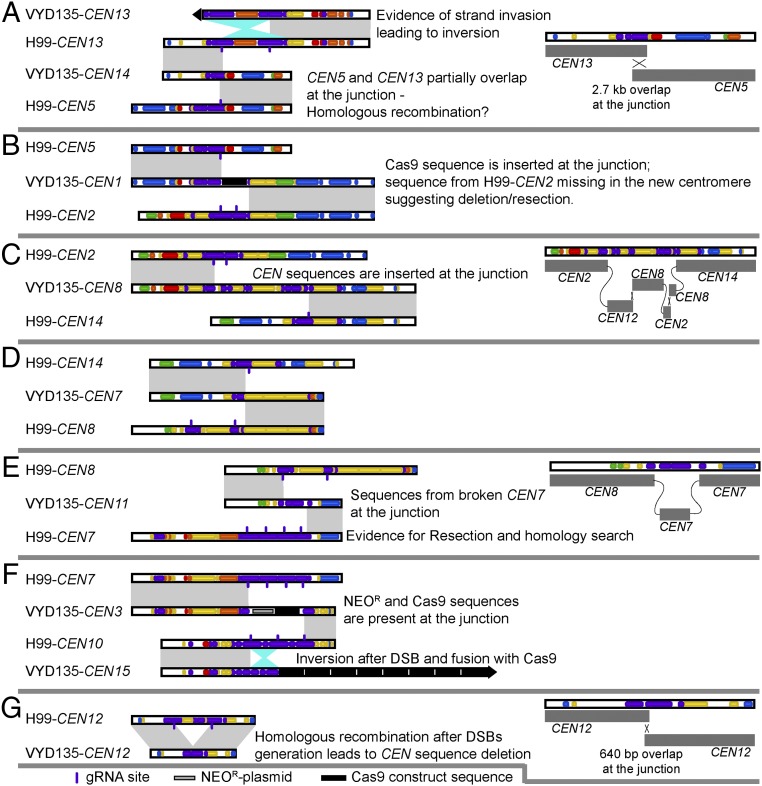
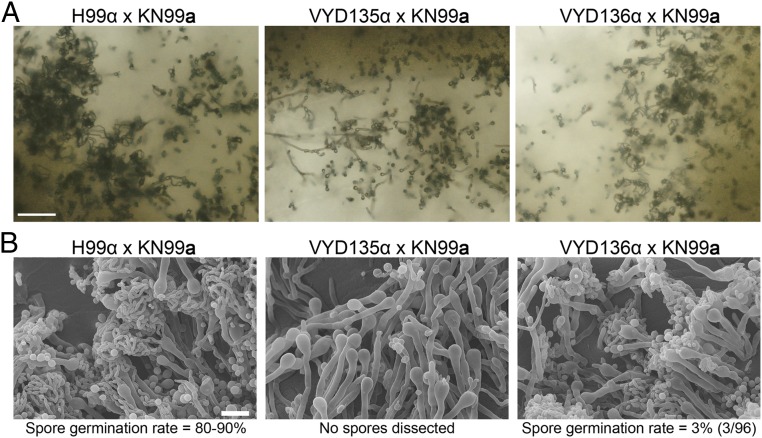
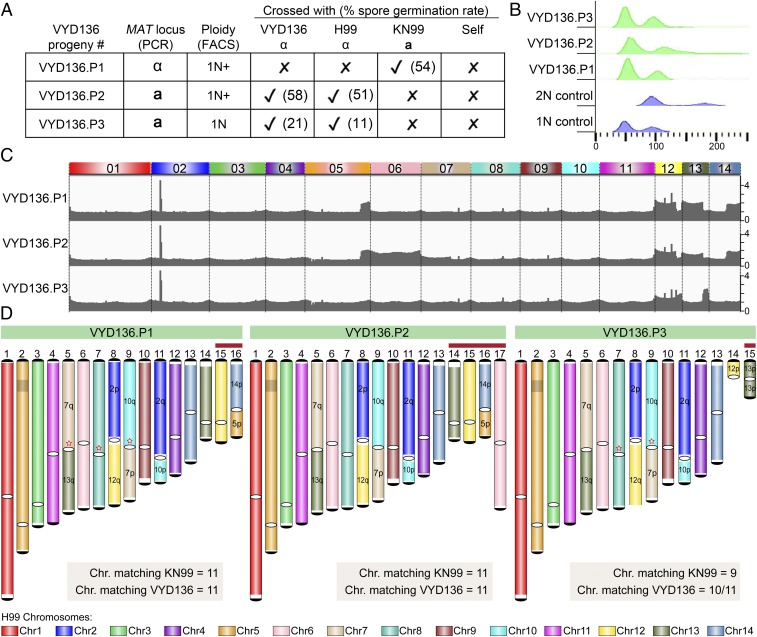
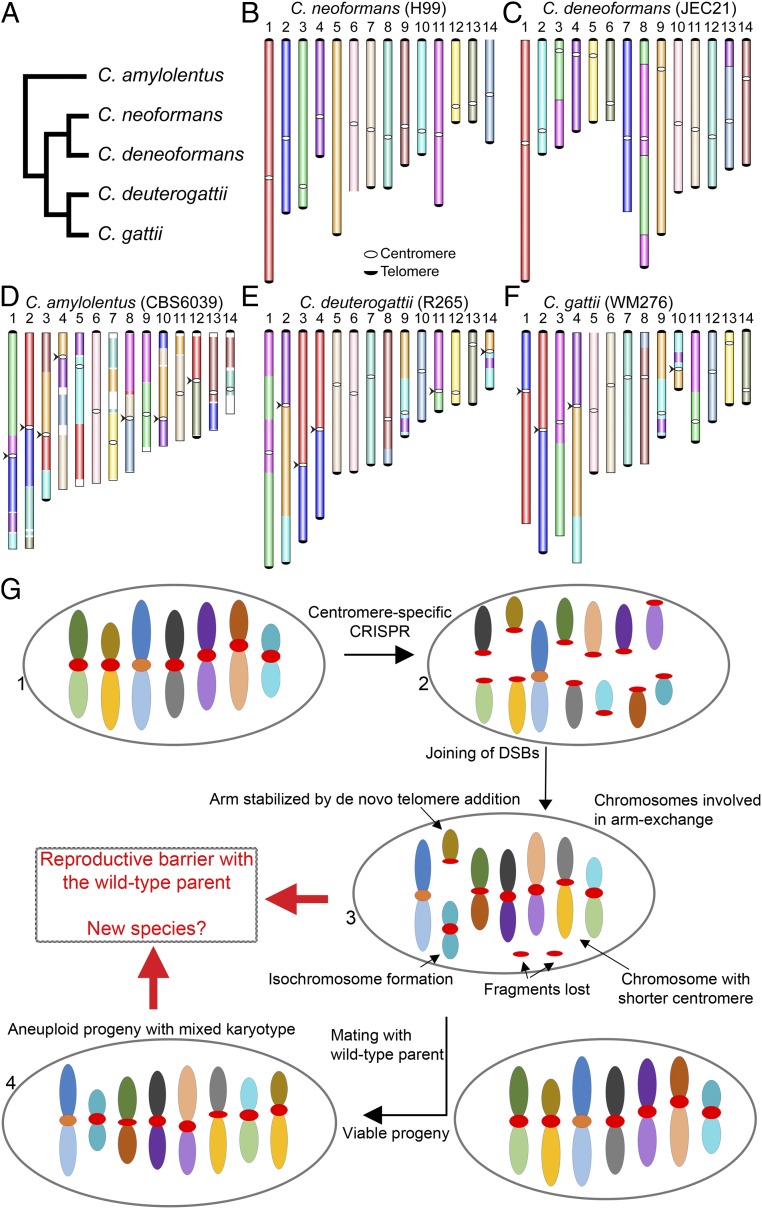
A fundamental characteristic of eukaryotic organisms is the generation of genetic variation via sexual reproduction. Conversely, significant large-scale genome structure variations could hamper sexual reproduction, causing reproductive isolation and promoting speciation. The underlying processes behind large-scale genome rearrangements are not well understood and include chromosome translocations involving centromeres. Recent genomic studies in the Cryptococcus species complex revealed that chromosome translocations generated via centromere recombination have reshaped the genomes of different species. In this study, multiple DNA double-strand breaks (DSBs) were generated via the CRISPR/Cas9 system at centromere-specific retrotransposons in the human fungal pathogen Cryptococcus neoformans The resulting DSBs were repaired in a complex manner, leading to the formation of multiple interchromosomal rearrangements and new telomeres, similar to chromothripsis-like events. The newly generated strains harboring chromosome translocations exhibited normal vegetative growth but failed to undergo successful sexual reproduction with the parental wild-type strain. One of these strains failed to produce any spores, while another produced ∼3% viable progeny. The germinated progeny exhibited aneuploidy for multiple chromosomes and showed improved fertility with both parents. All chromosome translocation events were accompanied without any detectable change in gene sequences and thus suggest that chromosomal translocations alone may play an underappreciated role in the onset of reproductive isolation and speciation.
Keywords: Cryptococcus neoformans; DSB repair; chromosome translocation; karyotype evolution; retrotransposons.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Harewood L., Fraser P., The impact of chromosomal rearrangements on regulation of gene expression. Hum. Mol. Genet. 23, R76–R82 (2014). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
