

Autophagy promotes mammalian survival by suppressing oxidative stress and p53
- PMID: 32193353
- PMCID: PMC7197357
- DOI: 10.1101/gad.335570.119
Autophagy promotes mammalian survival by suppressing oxidative stress and p53
Abstract
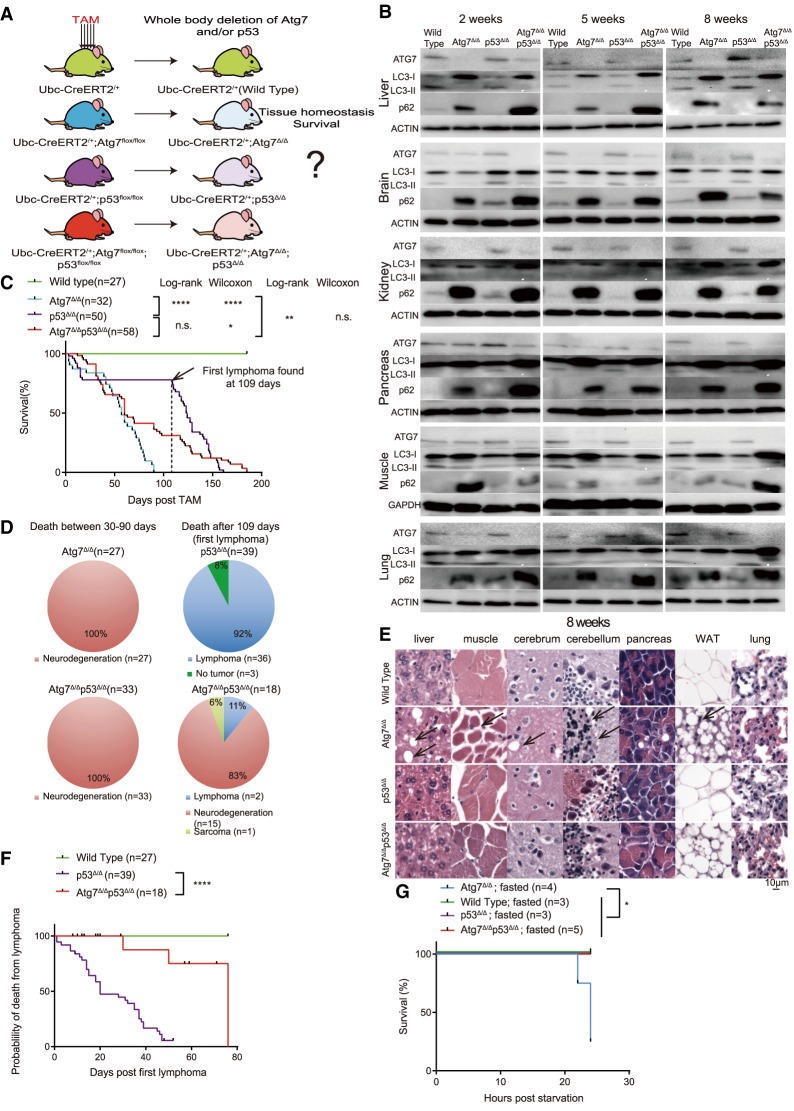
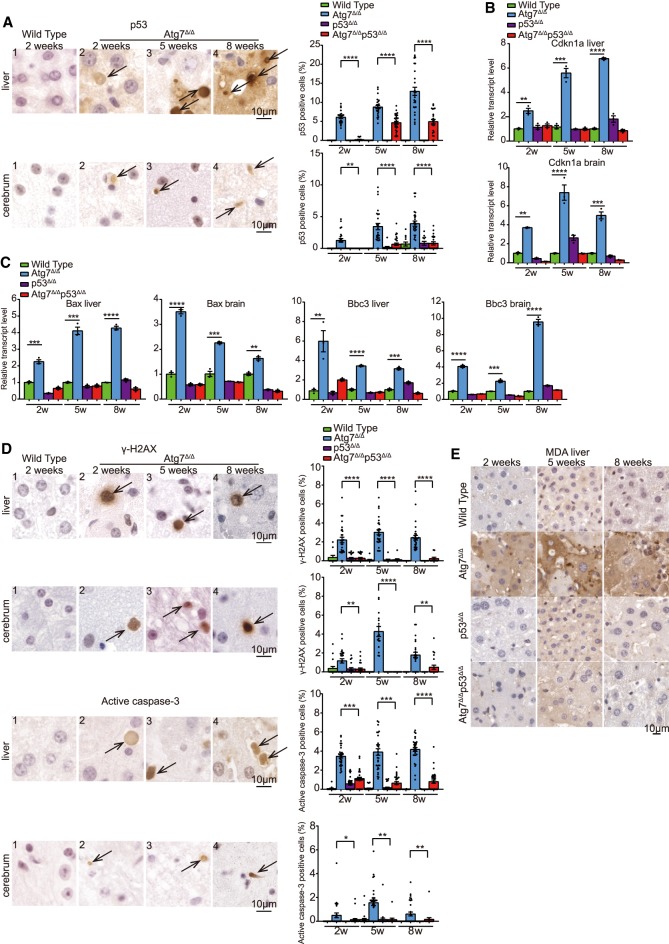
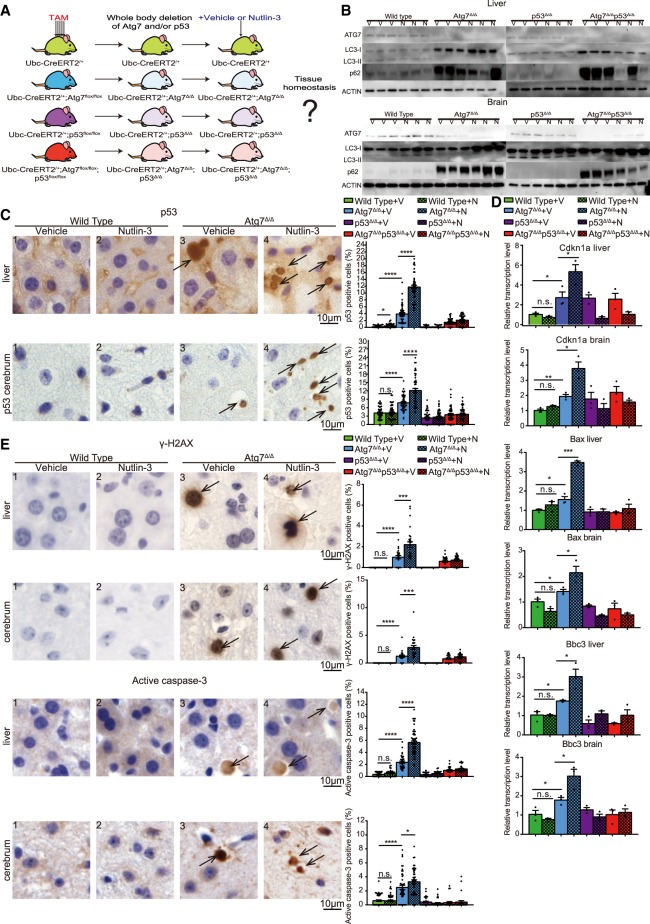
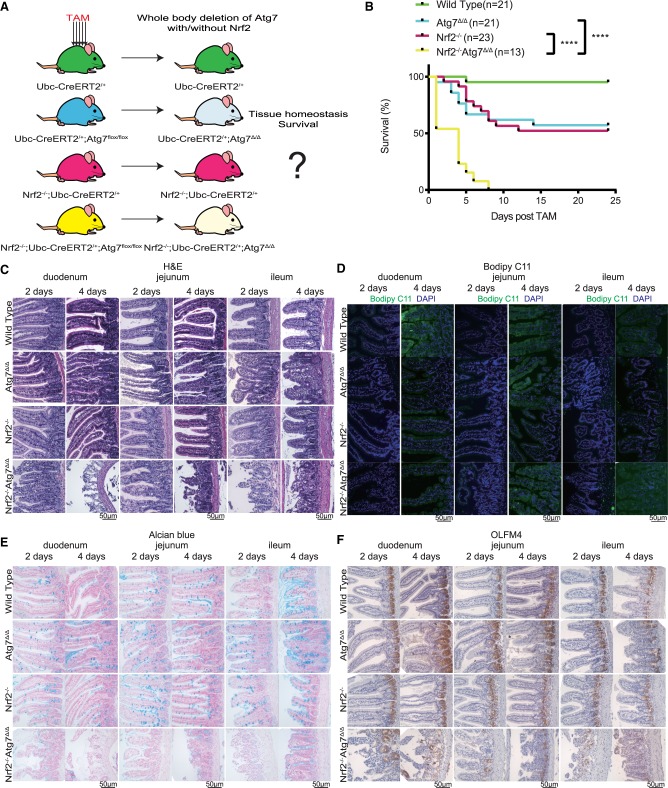
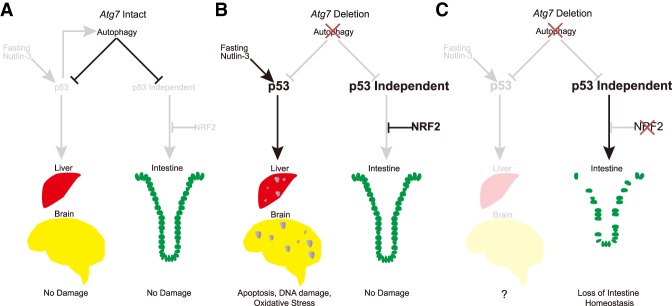
Autophagy captures intracellular components and delivers them to lysosomes for degradation and recycling. Conditional autophagy deficiency in adult mice causes liver damage, shortens life span to 3 mo due to neurodegeneration, and is lethal upon fasting. As autophagy deficiency causes p53 induction and cell death in neurons, we sought to test whether p53 mediates the lethal consequences of autophagy deficiency. Here, we conditionally deleted Trp53 (p53 hereafter) and/or the essential autophagy gene Atg7 throughout adult mice. Compared with Atg7Δ/Δ mice, the life span of Atg7Δ/Δp53Δ/Δ mice was extended due to delayed neurodegeneration and resistance to death upon fasting. Atg7 also suppressed apoptosis induced by p53 activator Nutlin-3, suggesting that autophagy inhibited p53 activation. To test whether increased oxidative stress in Atg7Δ/Δ mice was responsible for p53 activation, Atg7 was deleted in the presence or absence of the master regulator of antioxidant defense nuclear factor erythroid 2-related factor 2 (Nrf2). Nrf2-/-Atg7Δ/Δ mice died rapidly due to small intestine damage, which was not rescued by p53 codeletion. Thus, Atg7 limits p53 activation and p53-mediated neurodegeneration. In turn, NRF2 mitigates lethal intestine degeneration upon autophagy loss. These findings illustrate the tissue-specific roles for autophagy and functional dependencies on the p53 and NRF2 stress response mechanisms.
Keywords: ATG7; DNA damage; NRF2; apoptosis; autophagy; oxidative stress; p53.
© 2020 Yang et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Cox DR, Oakes D. 1984. Analysis of survival data. Chapman and Hall, London; New York.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous