

Endoplasmic reticulum-associated degradation regulates mitochondrial dynamics in brown adipocytes
- PMID: 32193362
- PMCID: PMC7409365
- DOI: 10.1126/science.aay2494
Endoplasmic reticulum-associated degradation regulates mitochondrial dynamics in brown adipocytes
Abstract
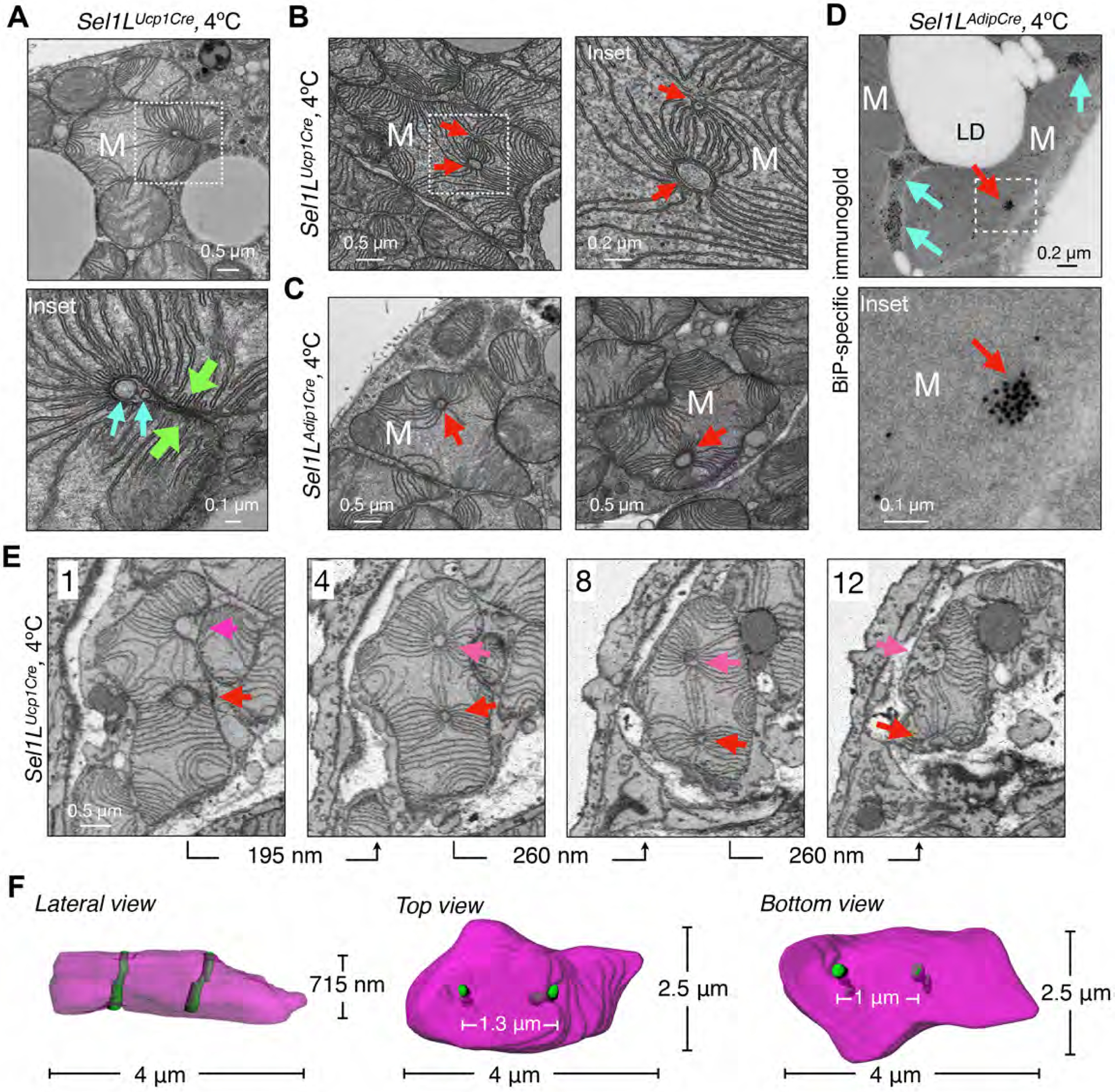
The endoplasmic reticulum (ER) engages mitochondria at specialized ER domains known as mitochondria-associated membranes (MAMs). Here, we used three-dimensional high-resolution imaging to investigate the formation of pleomorphic "megamitochondria" with altered MAMs in brown adipocytes lacking the Sel1L-Hrd1 protein complex of ER-associated protein degradation (ERAD). Mice with ERAD deficiency in brown adipocytes were cold sensitive and exhibited mitochondrial dysfunction. ERAD deficiency affected ER-mitochondria contacts and mitochondrial dynamics, at least in part, by regulating the turnover of the MAM protein, sigma receptor 1 (SigmaR1). Thus, our study provides molecular insights into ER-mitochondrial cross-talk and expands our understanding of the physiological importance of Sel1L-Hrd1 ERAD.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Competing interests:
The authors declare no conflict of interest.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
- UL1 TR000433/TR/NCATS NIH HHS/United States
- R01 DK107583/DK/NIDDK NIH HHS/United States
- R01 NS038619/NS/NINDS NIH HHS/United States
- R01 NS091242/NS/NINDS NIH HHS/United States
- R35 GM130292/GM/NIGMS NIH HHS/United States
- F32 CA228328/CA/NCI NIH HHS/United States
- R01 DK120047/DK/NIDDK NIH HHS/United States
- R01 GM113188/GM/NIGMS NIH HHS/United States
- P30 DK034933/DK/NIDDK NIH HHS/United States
- R01 GM123266/GM/NIGMS NIH HHS/United States
- P30 DK089503/DK/NIDDK NIH HHS/United States
- R01 GM130695/GM/NIGMS NIH HHS/United States
- R01 DK105393/DK/NIDDK NIH HHS/United States
- P20 GM121176/GM/NIGMS NIH HHS/United States
- R01 NS076715/NS/NINDS NIH HHS/United States
- P30 AR069620/AR/NIAMS NIH HHS/United States
- R01 DK110439/DK/NIDDK NIH HHS/United States
- L30 CA231806/CA/NCI NIH HHS/United States
- K99 CA241357/CA/NCI NIH HHS/United States
- R01 HL144657/HL/NHLBI NIH HHS/United States
- U24 DK097153/DK/NIDDK NIH HHS/United States
- R01 DK120330/DK/NIDDK NIH HHS/United States
