

GLP-1 Notch-LAG-1 CSL control of the germline stem cell fate is mediated by transcriptional targets lst-1 and sygl-1
- PMID: 32196486
- PMCID: PMC7153901
- DOI: 10.1371/journal.pgen.1008650
GLP-1 Notch-LAG-1 CSL control of the germline stem cell fate is mediated by transcriptional targets lst-1 and sygl-1
Abstract
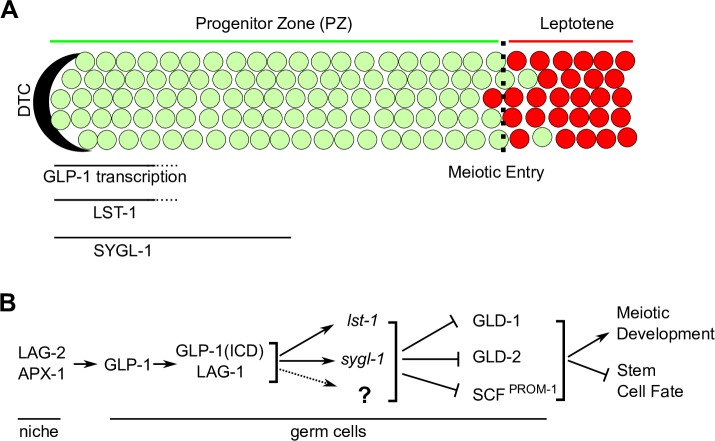
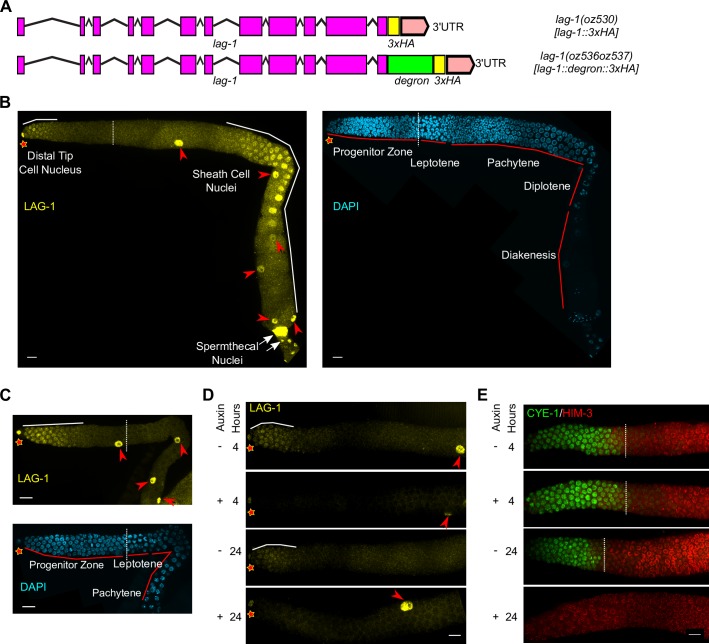
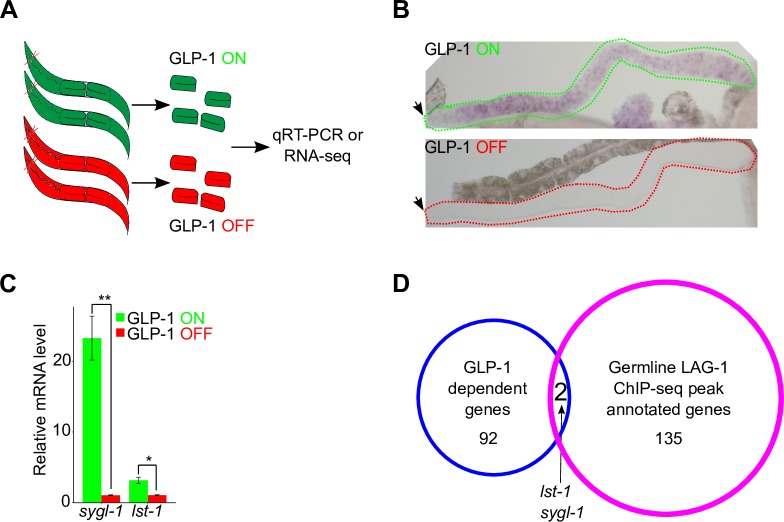
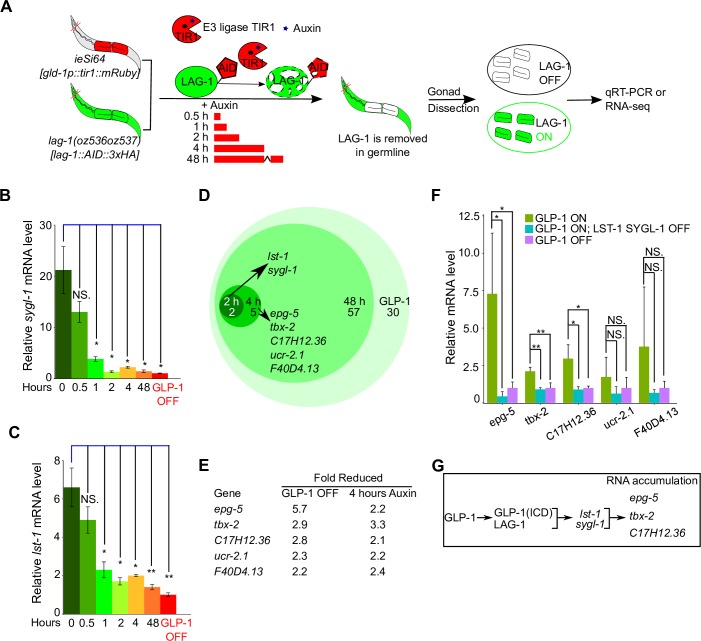
Stem cell systems are essential for the development and maintenance of polarized tissues. Intercellular signaling pathways control stem cell systems, where niche cells signal stem cells to maintain the stem cell fate/self-renewal and inhibit differentiation. In the C. elegans germline, GLP-1 Notch signaling specifies the stem cell fate, employing the sequence-specific DNA binding protein LAG-1 to implement the transcriptional response. We undertook a comprehensive genome-wide approach to identify transcriptional targets of GLP-1 signaling. We expected primary response target genes to be evident at the intersection of genes identified as directly bound by LAG-1, from ChIP-seq experiments, with genes identified as requiring GLP-1 signaling for RNA accumulation, from RNA-seq analysis. Furthermore, we performed a time-course transcriptomics analysis following auxin inducible degradation of LAG-1 to distinguish between genes whose RNA level was a primary or secondary response of GLP-1 signaling. Surprisingly, only lst-1 and sygl-1, the two known target genes of GLP-1 in the germline, fulfilled these criteria, indicating that these two genes are the primary response targets of GLP-1 Notch and may be the sole germline GLP-1 signaling protein-coding transcriptional targets for mediating the stem cell fate. In addition, three secondary response genes were identified based on their timing following loss of LAG-1, their lack of a LAG-1 ChIP-seq peak and that their glp-1 dependent mRNA accumulation could be explained by a requirement for lst-1 and sygl-1 activity. Moreover, our analysis also suggests that the function of the primary response genes lst-1 and sygl-1 can account for the glp-1 dependent peak protein accumulation of FBF-2, which promotes the stem cell fate and, in part, for the spatial restriction of elevated LAG-1 accumulation to the stem cell region.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
