

Autotrophic carbon fixation pathways along the redox gradient in oxygen-depleted oceanic waters
- PMID: 32202395
- PMCID: PMC7318340
- DOI: 10.1111/1758-2229.12837
Autotrophic carbon fixation pathways along the redox gradient in oxygen-depleted oceanic waters
Abstract
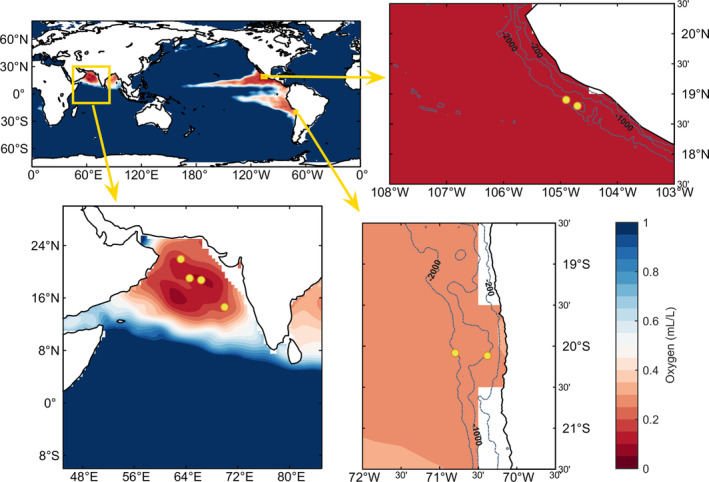
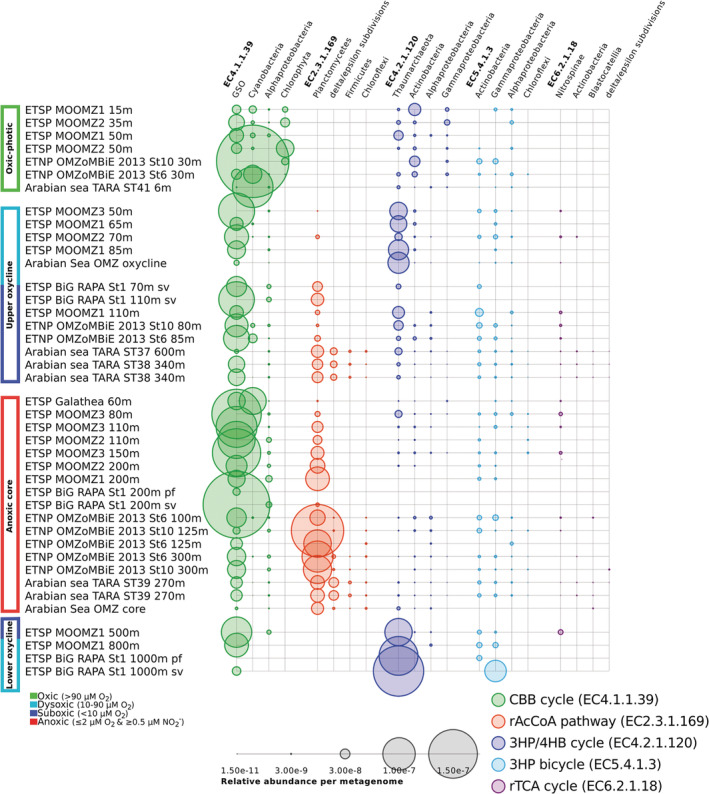
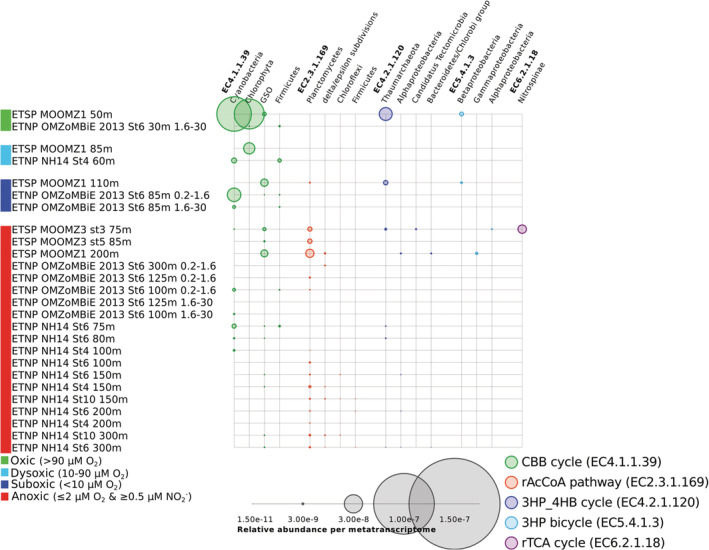
Anoxic marine zones (AMZs), also known as 'oxygen-deficient zones', contribute to the loss of fixed nitrogen from the ocean by anaerobic microbial processes. While these microbial processes associated with the nitrogen cycle have been extensively studied, those linked to the carbon cycle in AMZs have received much less attention, particularly the autotrophic carbon fixation - a crucial component of the carbon cycle. Using metagenomic and metatranscriptomic data from major AMZs, we report an explicit partitioning of the marker genes associated with different autotrophic carbon fixation pathways along the redox gradient (from oxic to anoxic conditions) present in the water column of AMZs. Sequences related to the Calvin-Benson-Bassham cycle were found along the entire gradient, while those related to the reductive Acetyl-CoA pathway were restricted to suboxic and anoxic waters. Sequences putatively associated with the 3-hydroxypropionate/4-hydroxybutyrate cycle dominated in the upper and lower oxyclines. Genes related to the reductive tricarboxylic acid cycle were represented from dysoxic to anoxic waters. The taxonomic affiliation of the sequences is consistent with the presence of microorganisms involved in crucial steps of biogeochemical cycles in AMZs, such as the gamma-proteobacteria sulfur oxidisers, the anammox bacteria Candidatus Scalindua and the thaumarcheota ammonia oxidisers of the Marine Group I.
© 2020 The Authors. Environmental Microbiology Reports published by Society for Applied Microbiology and John Wiley & Sons Ltd.
Figures
References
-
- Belmar, L. , Molina, V. , and Ulloa, O. (2011) Abundance and phylogenetic identity of archaeoplankton in the permanent oxygen minimum zone of the eastern tropical South Pacific. FEMS Microbiol Ecol 78: 314–326. - PubMed
-
- Berg, I.A. , Kockelkorn, D. , Ramos‐Vera, W.H. , Say, R.F. , Zarzycki, J. , Hügler, M. , et al (2010) Autotrophic carbon fixation in archaea. Nat Rev Microbiol 8: 447–460. - PubMed
-
- Canfield, D.E. , Stewart, F.J. , Thamdrup, B. , Brabandere, L.D. , Dalsgaard, T. , Delong, E.F. , et al (2010) A cryptic sulfur cycle in oxygen‐minimum‐zone waters off the Chilean coast. Science 330: 1375–1378. - PubMed
-
- Caspi, R. , Billington, R. , Fulcher, C.A. , Keseler, I.M. , Kothari, A. , Krummenacker, M. , et al (2017) The meta Cyc database of metabolic pathways and enzymes. Nucl Acids Res 46: D623–D631.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
