

Phase separation of TAZ compartmentalizes the transcription machinery to promote gene expression
- PMID: 32203417
- PMCID: PMC11044910
- DOI: 10.1038/s41556-020-0485-0
Phase separation of TAZ compartmentalizes the transcription machinery to promote gene expression
Abstract
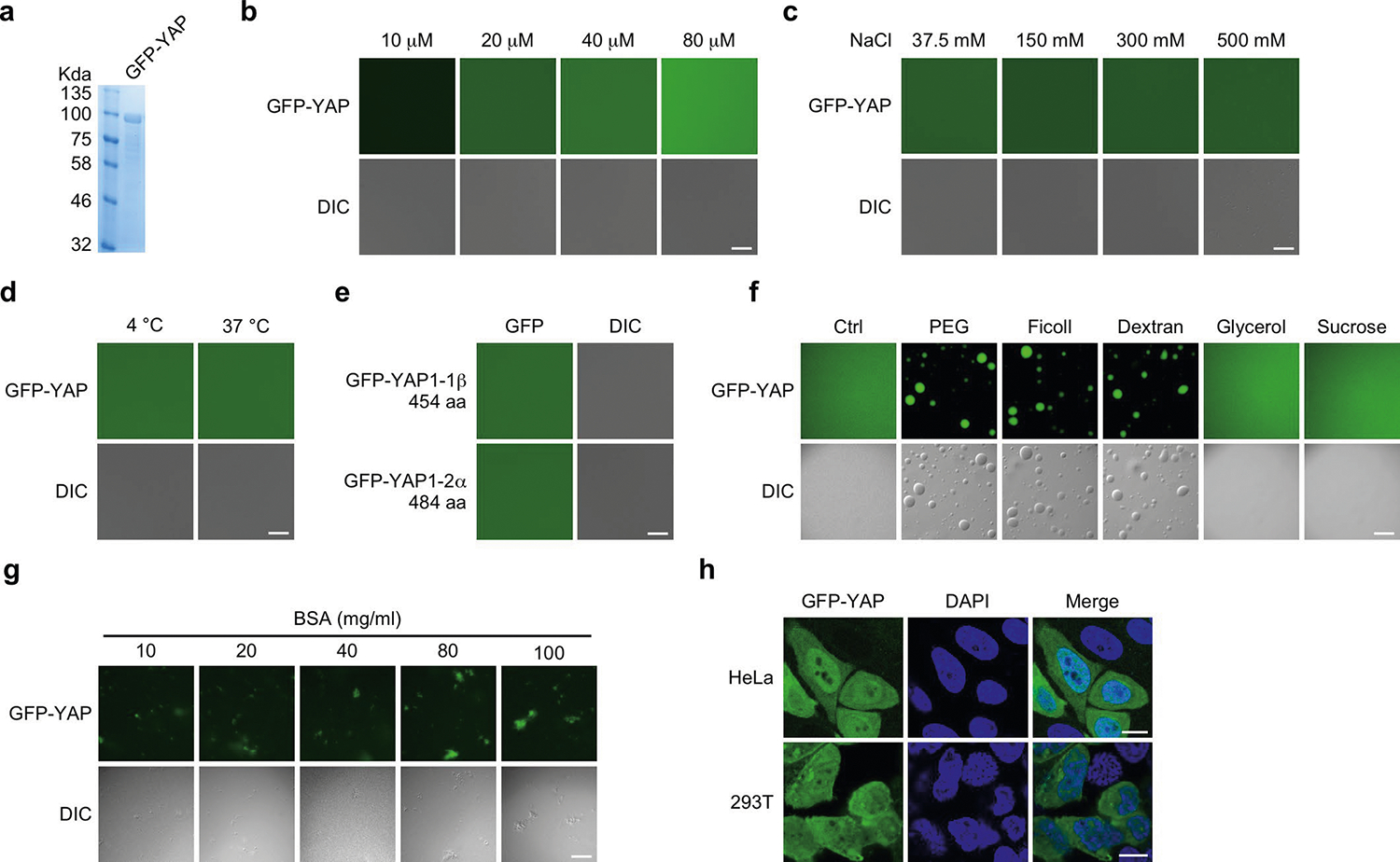
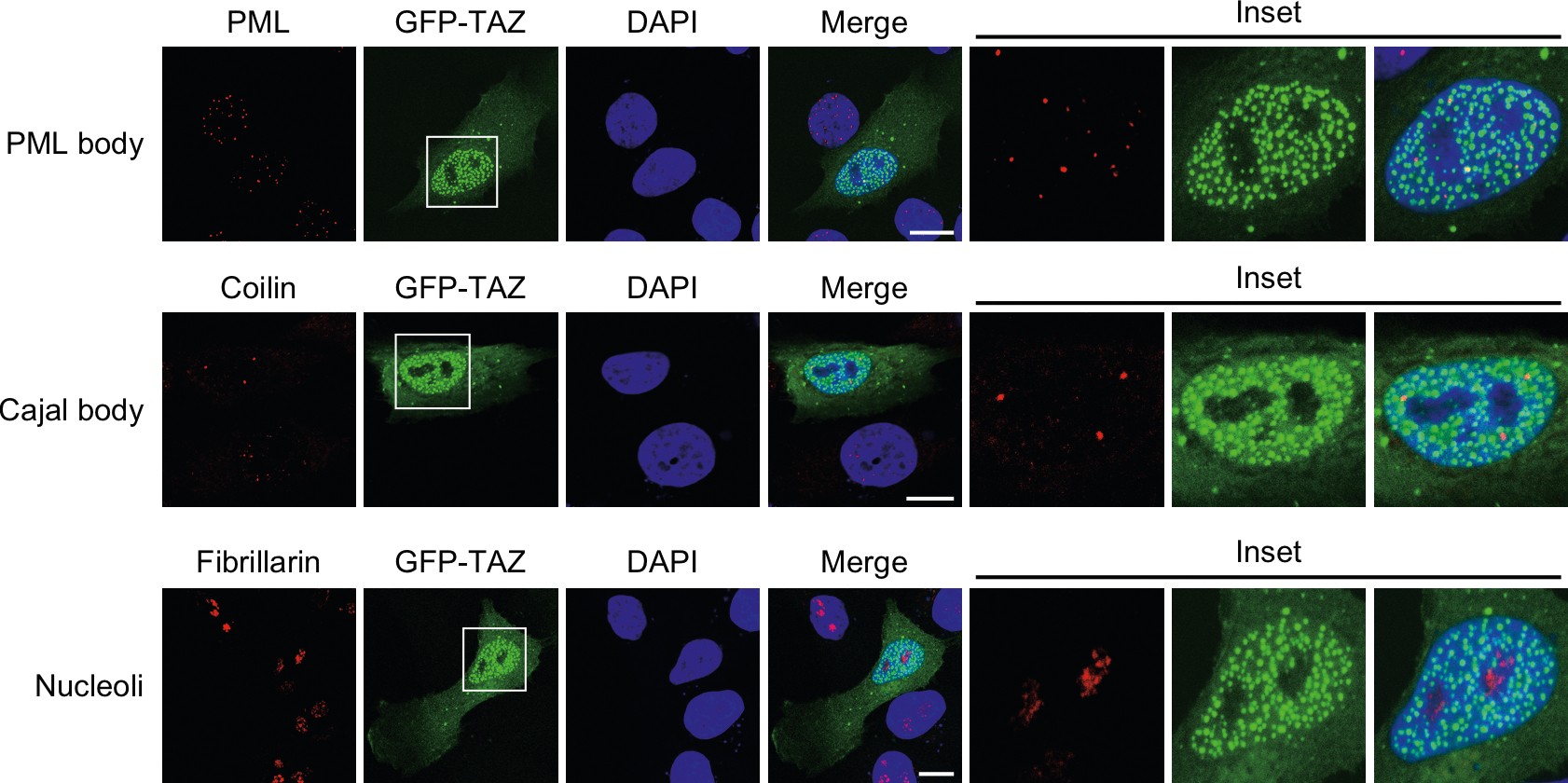
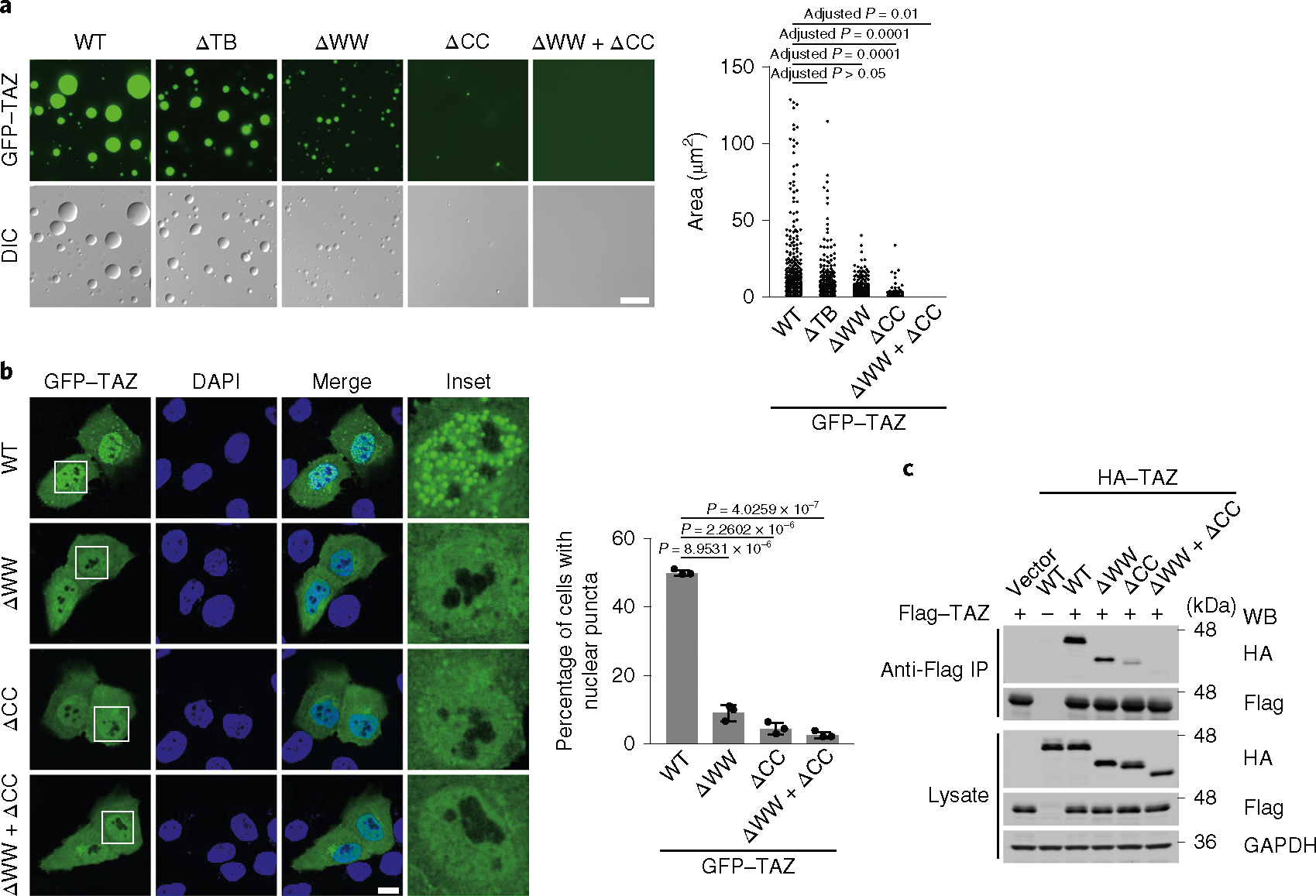
TAZ promotes growth, development and tumorigenesis by regulating the expression of target genes. However, the manner in which TAZ orchestrates the transcriptional responses is poorly defined. Here we demonstrate that TAZ forms nuclear condensates through liquid-liquid phase separation to compartmentalize its DNA-binding cofactor TEAD4, coactivators BRD4 and MED1, and the transcription elongation factor CDK9 for transcription. TAZ forms phase-separated droplets in vitro and liquid-like nuclear condensates in vivo, and this ability is negatively regulated by Hippo signalling through LATS-mediated phosphorylation and is mediated by the coiled-coil (CC) domain. Deletion of the TAZ CC domain or substitution with the YAP CC domain prevents the phase separation of TAZ and its ability to induce the expression of TAZ-specific target genes. Thus, we identify a mechanism of transcriptional activation by TAZ and demonstrate that pathway-specific transcription factors also engage the phase-separation mechanism for efficient and specific transcriptional activation.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures













Comment in
-
YAP/TAZ phase separation for transcription.Nat Cell Biol. 2020 Apr;22(4):357-358. doi: 10.1038/s41556-020-0498-8. Nat Cell Biol. 2020. PMID: 32203419 No abstract available.
References
-
- Ma S, Meng Z, Chen R & Guan KL The Hippo pathway: biology and pathophysiology. Annu. Rev. Biochem. 88, 577–604 (2018). - PubMed
-
- Tapon N & Harvey KF The Hippo pathway—from top to bottom and everything in between. Semin. Cell Dev. Biol. 23, 768–769 (2012). - PubMed
-
- Badouel C & McNeill H SnapShot: the Hippo signaling pathway. Cell 145, 484 (2011). - PubMed
-
- Schroeder MC & Halder G Regulation of the Hippo pathway by cell architecture and mechanical signals. Semin. Cell Dev. Biol. 23, 803–811 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
