

A neuroglobin-based high-affinity ligand trap reverses carbon monoxide-induced mitochondrial poisoning
- PMID: 32205448
- PMCID: PMC7212636
- DOI: 10.1074/jbc.RA119.010593
A neuroglobin-based high-affinity ligand trap reverses carbon monoxide-induced mitochondrial poisoning
Abstract
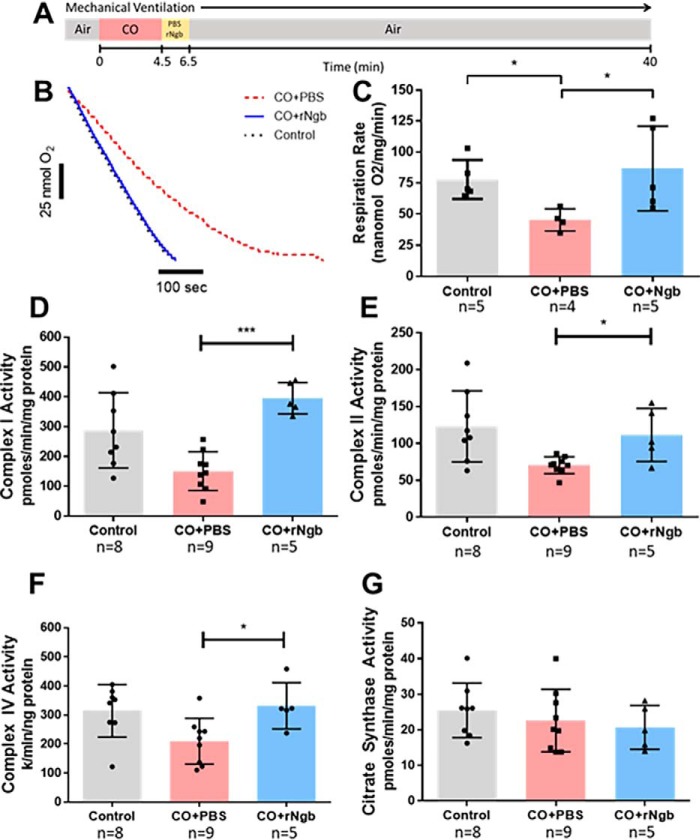
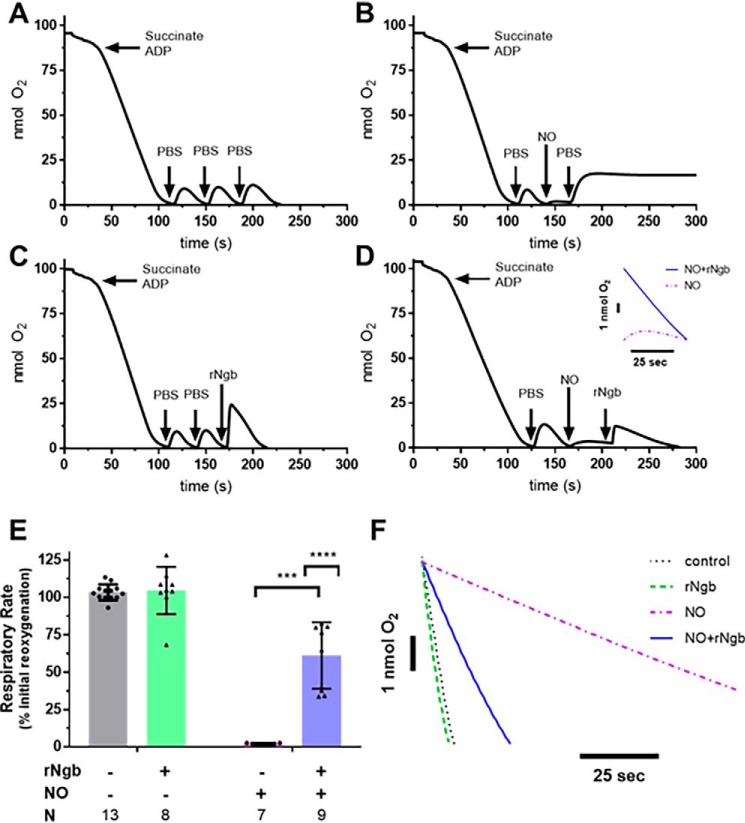
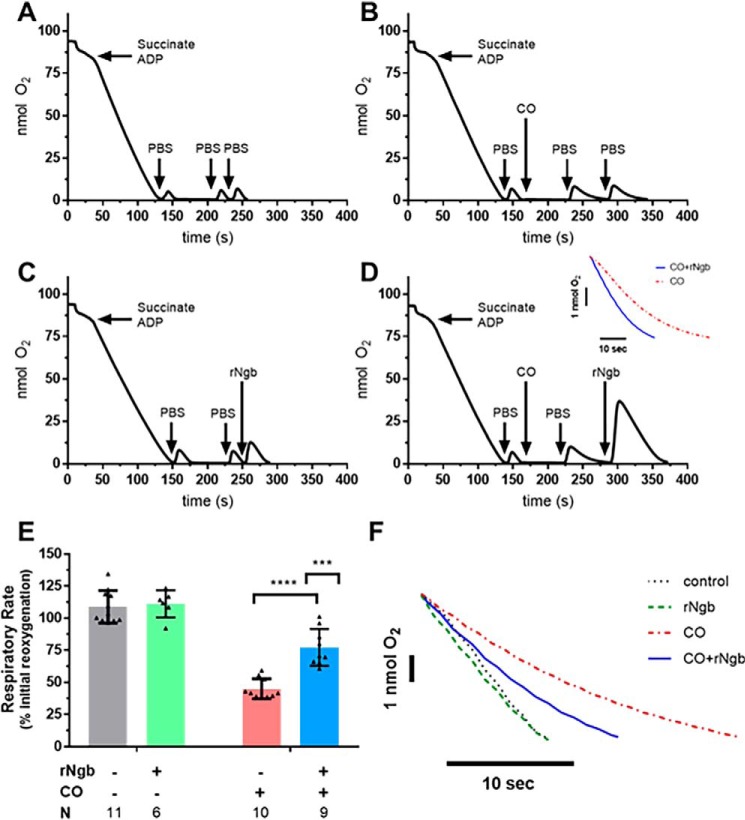
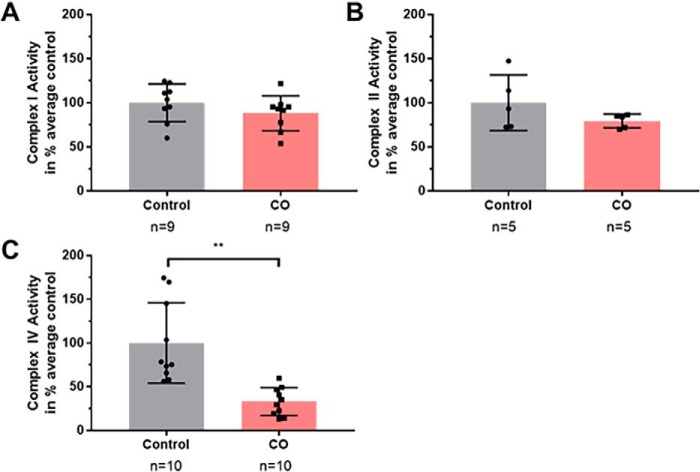
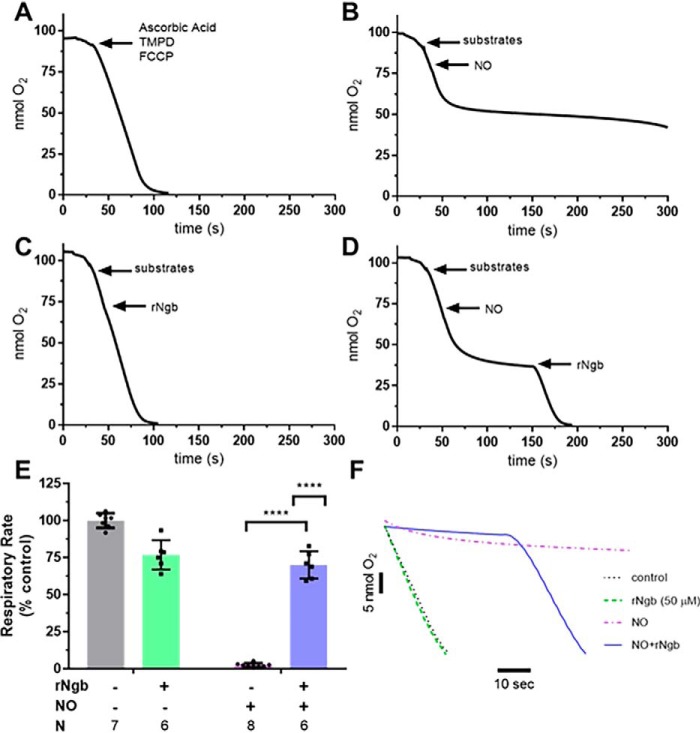
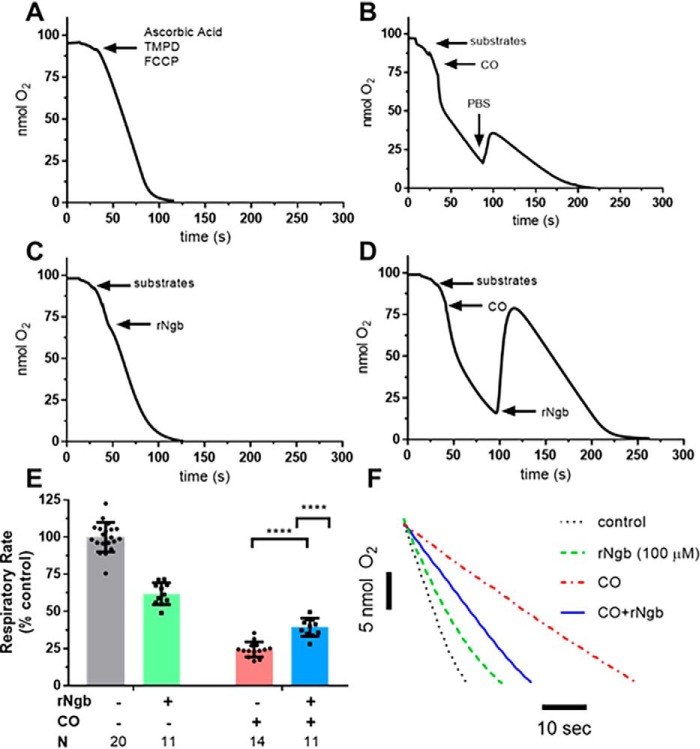
Carbon monoxide (CO) remains the most common cause of human poisoning. The consequences of CO poisoning include cardiac dysfunction, brain injury, and death. CO causes toxicity by binding to hemoglobin and by inhibiting mitochondrial cytochrome c oxidase (CcO), thereby decreasing oxygen delivery and inhibiting oxidative phosphorylation. We have recently developed a CO antidote based on human neuroglobin (Ngb-H64Q-CCC). This molecule enhances clearance of CO from red blood cells in vitro and in vivo Herein, we tested whether Ngb-H64Q-CCC can also scavenge CO from CcO and attenuate CO-induced inhibition of mitochondrial respiration. Heart tissue from mice exposed to 3% CO exhibited a 42 ± 19% reduction in tissue respiration rate and a 33 ± 38% reduction in CcO activity compared with unexposed mice. Intravenous infusion of Ngb-H64Q-CCC restored respiration rates to that of control mice correlating with higher electron transport chain CcO activity in Ngb-H64Q-CCC-treated compared with PBS-treated, CO-poisoned mice. Further, using a Clark-type oxygen electrode, we measured isolated rat liver mitochondrial respiration in the presence and absence of saturating solutions of CO (160 μm) and nitric oxide (100 μm). Both CO and NO inhibited respiration, and treatment with Ngb-H64Q-CCC (100 and 50 μm, respectively) significantly reversed this inhibition. These results suggest that Ngb-H64Q-CCC mitigates CO toxicity by scavenging CO from carboxyhemoglobin, improving systemic oxygen delivery and reversing the inhibitory effects of CO on mitochondria. We conclude that Ngb-H64Q-CCC or other CO scavengers demonstrate potential as antidotes that reverse the clinical and molecular effects of CO poisoning.
Keywords: CO poisoning; antidotes; carbon monoxide; hemoglobin; hypoxia; hypoxia-inducible factor (HIF); medical toxicology; mitochondria; mitochondrial disease; mitochondrial respiratory chain complex; neuroglobin; nitric oxide.
© 2020 Rose et al.
Conflict of interest statement
J. J. R., L. W., C. F. M., J. T., and M. T. G. are shareholders in Globin Solutions. J. J. R., Q. X., A. W. D., J. T., and M. T. G. are coinventors of provisional, pending, and granted patents for the use of recombinant neuroglobin and other heme-based molecules as antidotes for carbon monoxide poisoning. J. J. R. and J. T. are officers and directors of Globin Solutions, Inc. M. T. G. is a director and advisor of Globin Solutions, Inc., which has licensed this technology and had an option to technology directed at using hydroxycobalamin for carbon monoxide poisoning from Virginia Commonwealth University that has expired. M. T. G. is a coinventor on patents directed to the use of nitrite salts in cardiovascular diseases licensed and exclusively optioned to Globin Solutions, Inc., which has a sponsored research agreement with the University of Pittsburgh aimed at developing carbon monoxide poisoning antidotes into therapeutics that did not support the research contained in this grant that partially supports the efforts of M. T. G., J. T., Q. X., X. C., Q. T., and X. N. H., M. T. G. is a coinvestigator in a research collaboration with Bayer Pharmaceuticals to evaluate riociguate as a treatment for patients with sickle cell disease. The financial conflicts of interest of J. J. R., J. T., Q. X., L. W., A. W. D., C. F. M., and M. T. G. were managed by the University of Pittsburgh Conflict of Interest Committee and a data stewardship committee
Figures
References
-
- Hampson N. B. (2016) U.S. mortality due to carbon monoxide poisoning, 1999–2014: accidental and intentional deaths. Ann. Am. Thorac. Soc. 13, 1768–1774 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
