

The fruitless gene affects female receptivity and species isolation
- PMID: 32208837
- PMCID: PMC7126032
- DOI: 10.1098/rspb.2019.2765
The fruitless gene affects female receptivity and species isolation
Abstract
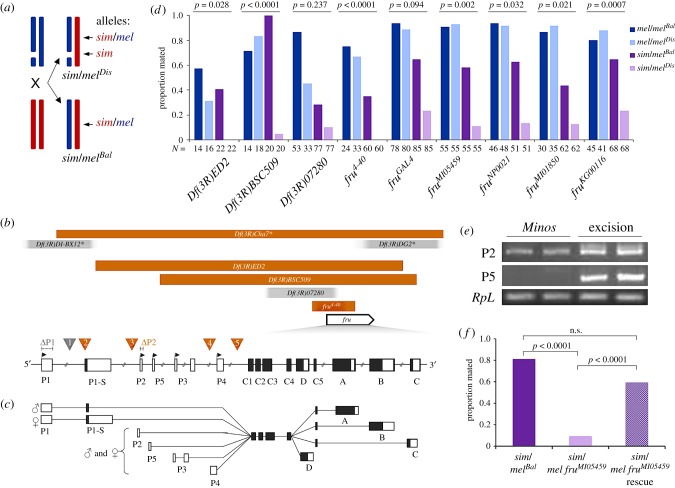
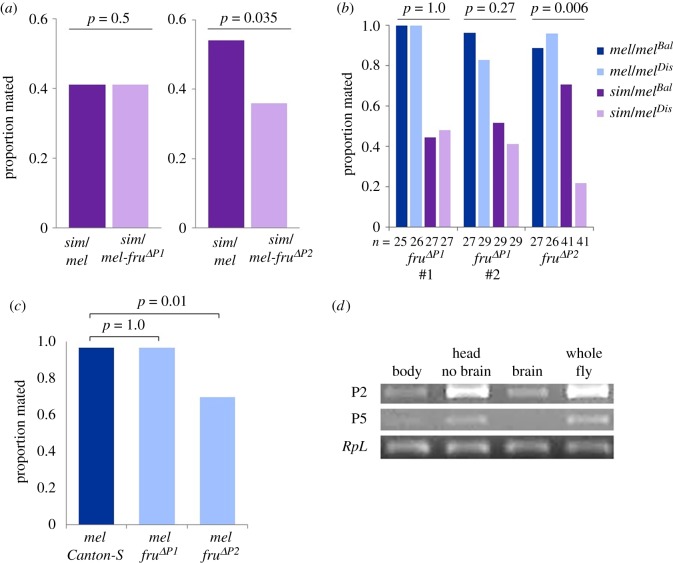
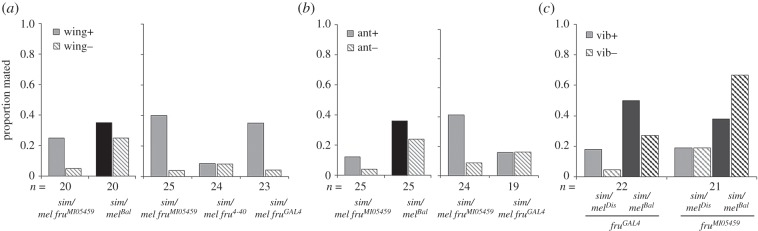
Female mate rejection acts as a major selective force within species, and can serve as a reproductive barrier between species. In spite of its critical role in fitness and reproduction, surprisingly little is known about the genetic or neural basis of variation in female mate choice. Here, we identify fruitless as a gene affecting female receptivity within Drosophila melanogaster, as well as female Drosophila simulans rejection of male D. melanogaster. Of the multiple transcripts this gene produces, by far the most widely studied is the sex-specifically spliced transcript involved in the sex determination pathway. However, we find that female rejection behaviour is affected by a non-sex-specifically spliced fruitless transcript. This is the first implication of fruitless in female behaviour, and the first behavioural role identified for a fruitless non-sex-specifically spliced transcript. We found that this locus does not influence preferences via a single sensory modality, examining courtship song, antennal pheromone perception, or perception of substrate vibrations, and we conclude that fruitless influences mate choice via the integration of multiple signals or through another sensory modality.
Keywords: Drosophila melanogaster; Drosophila simulans; behavioural isolation; fruitless; prezygotic isolation; speciation.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Trivers RL. 1972. Parental investment and sexual selection. In Sexual selection and the descent of man (ed. Campbell B.), pp. 136–179. Chicago, IL: Aldine Press.
-
- Parker G. 1983. Mate quality and mating decisions. In Mate choice (ed. Bateson PG.), pp. 141–166. Cambridge, UK: Cambridge University Press.
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
