

Lipid Anchoring of Archaeosortase Substrates and Midcell Growth in Haloarchaea
- PMID: 32209681
- PMCID: PMC7157517
- DOI: 10.1128/mBio.00349-20
Lipid Anchoring of Archaeosortase Substrates and Midcell Growth in Haloarchaea
Abstract
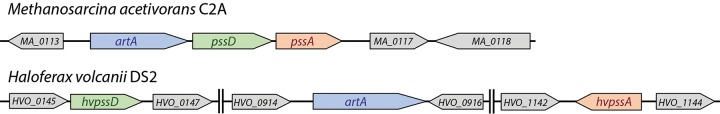
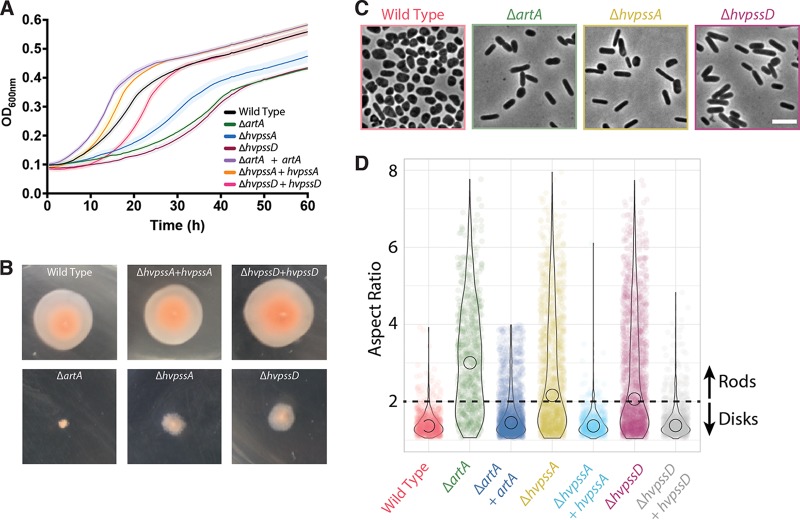
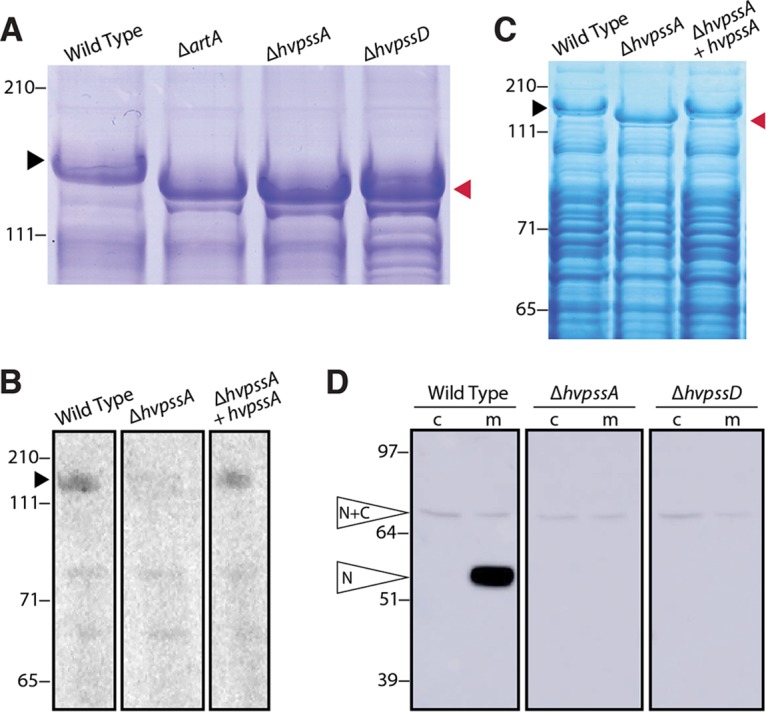
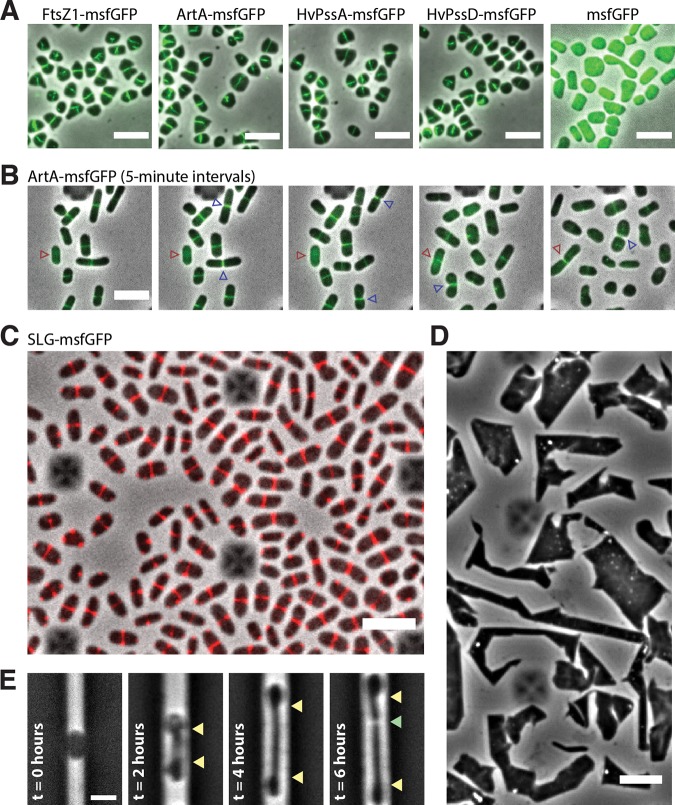
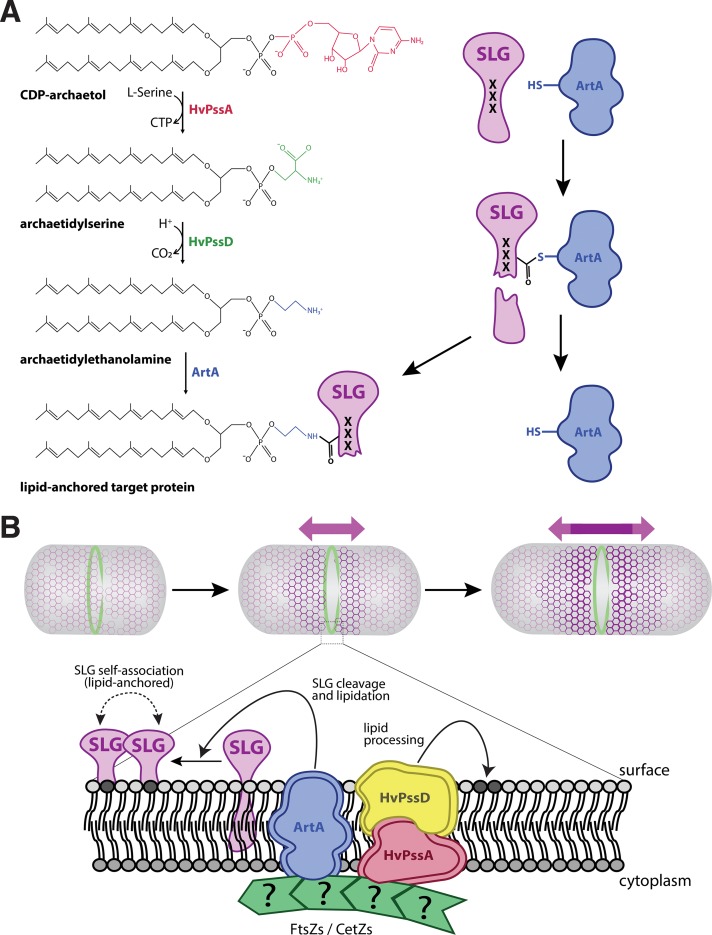
The archaeal cytoplasmic membrane provides an anchor for many surface proteins. Recently, a novel membrane anchoring mechanism involving a peptidase, archaeosortase A (ArtA), and C-terminal lipid attachment of surface proteins was identified in the model archaeon Haloferax volcanii ArtA is required for optimal cell growth and morphogenesis, and the S-layer glycoprotein (SLG), the sole component of the H. volcanii cell wall, is one of the targets for this anchoring mechanism. However, how exactly ArtA function and regulation control cell growth and morphogenesis is still elusive. Here, we report that archaeal homologs to the bacterial phosphatidylserine synthase (PssA) and phosphatidylserine decarboxylase (PssD) are involved in ArtA-dependent protein maturation. Haloferax volcanii strains lacking either HvPssA or HvPssD exhibited motility, growth, and morphological phenotypes similar to those of an ΔartA mutant. Moreover, we showed a loss of covalent lipid attachment to SLG in the ΔhvpssA mutant and that proteolytic cleavage of the ArtA substrate HVO_0405 was blocked in the ΔhvpssA and ΔhvpssD mutant strains. Strikingly, ArtA, HvPssA, and HvPssD green fluorescent protein (GFP) fusions colocalized to the midcell position of H. volcanii cells, strongly supporting that they are involved in the same pathway. Finally, we have shown that the SLG is also recruited to the midcell before being secreted and lipid anchored at the cell outer surface. Collectively, our data suggest that haloarchaea use the midcell as the main surface processing hot spot for cell elongation, division, and shape determination.IMPORTANCE The subcellular organization of biochemical processes in space and time is still one of the most mysterious topics in archaeal cell biology. Despite the fact that haloarchaea largely rely on covalent lipid anchoring to coat the cell envelope, little is known about how cells coordinate de novo synthesis and about the insertion of this proteinaceous layer throughout the cell cycle. Here, we report the identification of two novel contributors to ArtA-dependent lipid-mediated protein anchoring to the cell surface, HvPssA and HvPssD. ArtA, HvPssA, and HvPssD, as well as SLG, showed midcell localization during growth and cytokinesis, indicating that haloarchaeal cells confine phospholipid processing in order to promote midcell elongation. Our findings have important implications for the biogenesis of the cell surface.
Keywords: Haloferax volcanii; S-layer; archaea; archaeosortase; cell division; cell elongation; cell shape; cell surface; haloarchaea; lipid anchoring.
Copyright © 2020 Abdul-Halim et al.
Figures
References
-
- May JW, Mitchison JM. 1986. Length growth in fission yeast cells measured by two novel techniques. Nature 322:752–754. doi: 10.1038/322752a0. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
