

Sperm SPACA6 protein is required for mammalian Sperm-Egg Adhesion/Fusion
- PMID: 32210282
- PMCID: PMC7093486
- DOI: 10.1038/s41598-020-62091-y
Sperm SPACA6 protein is required for mammalian Sperm-Egg Adhesion/Fusion
Abstract
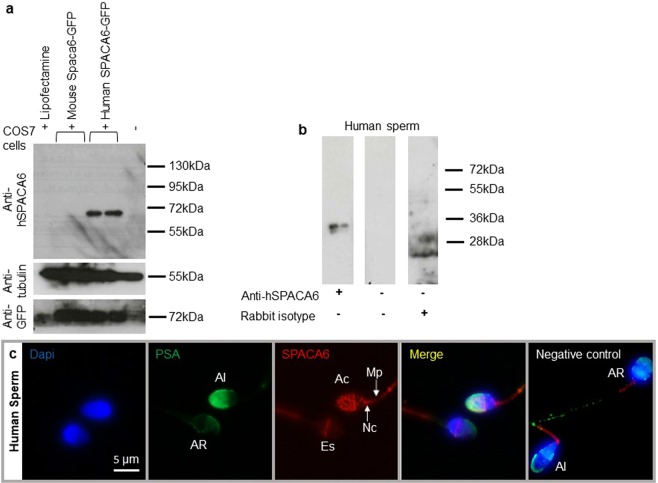
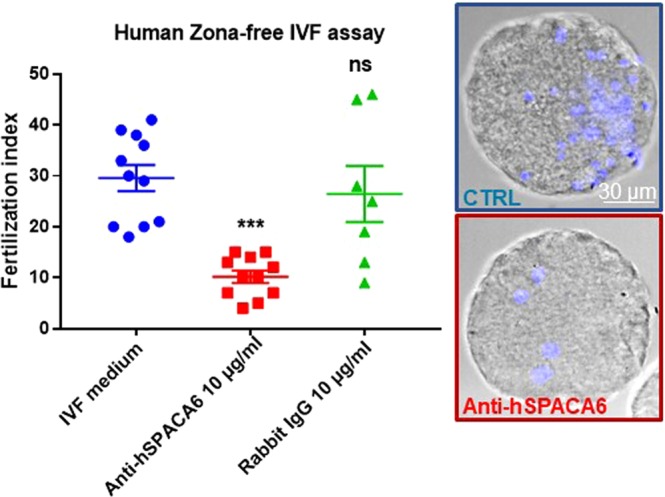
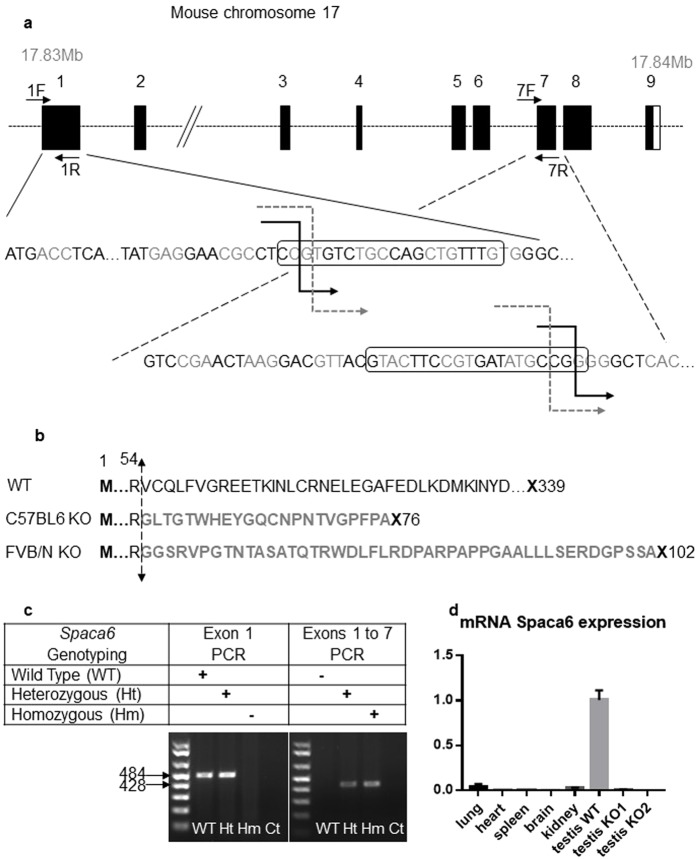
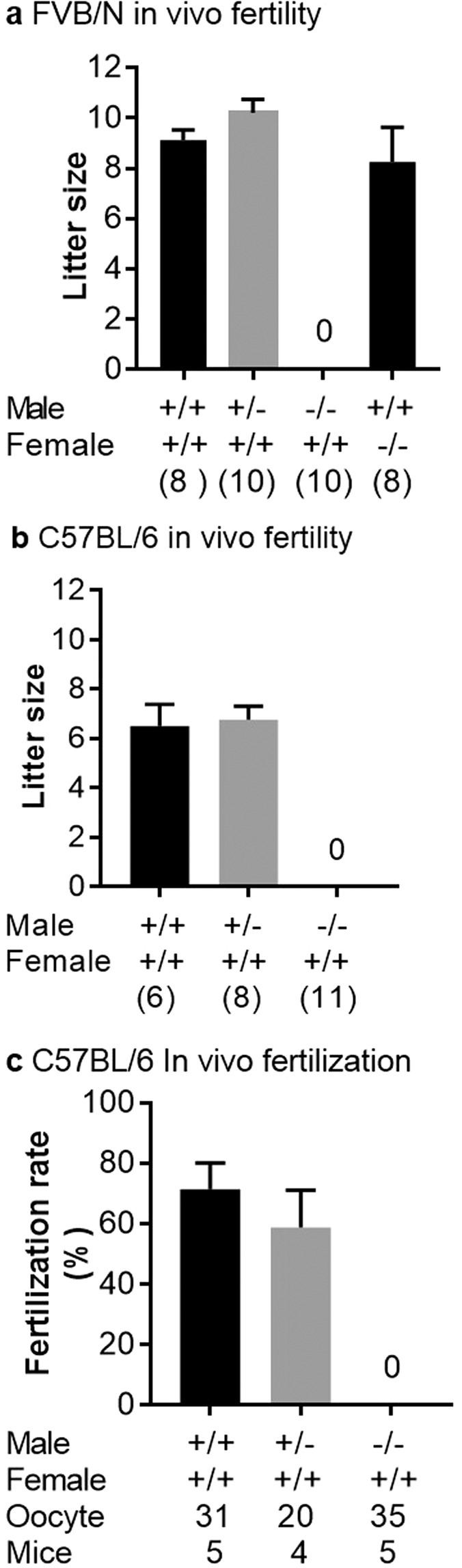
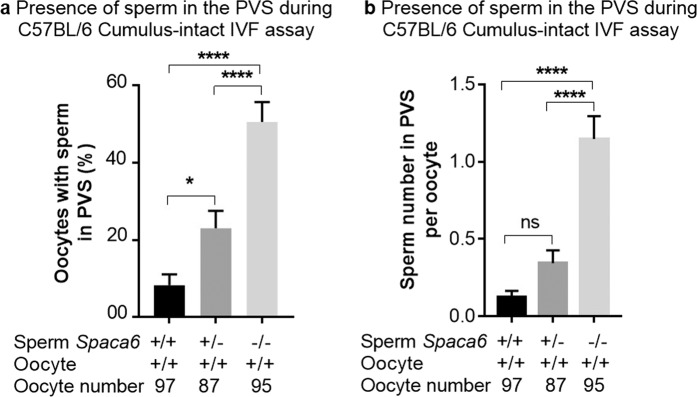
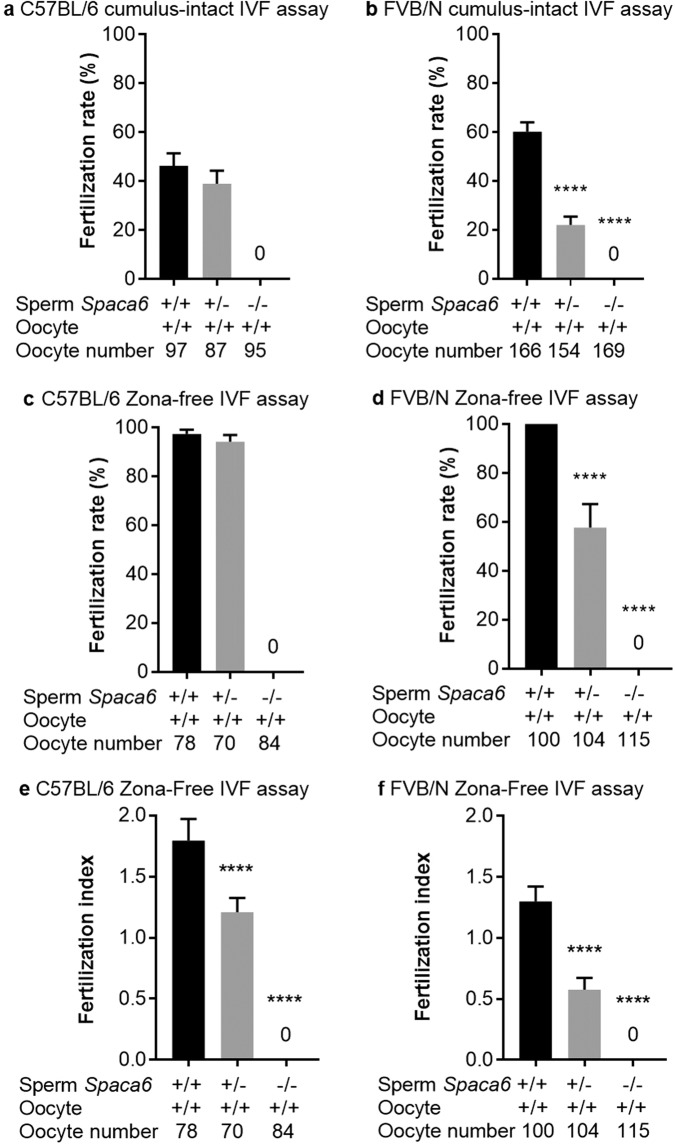
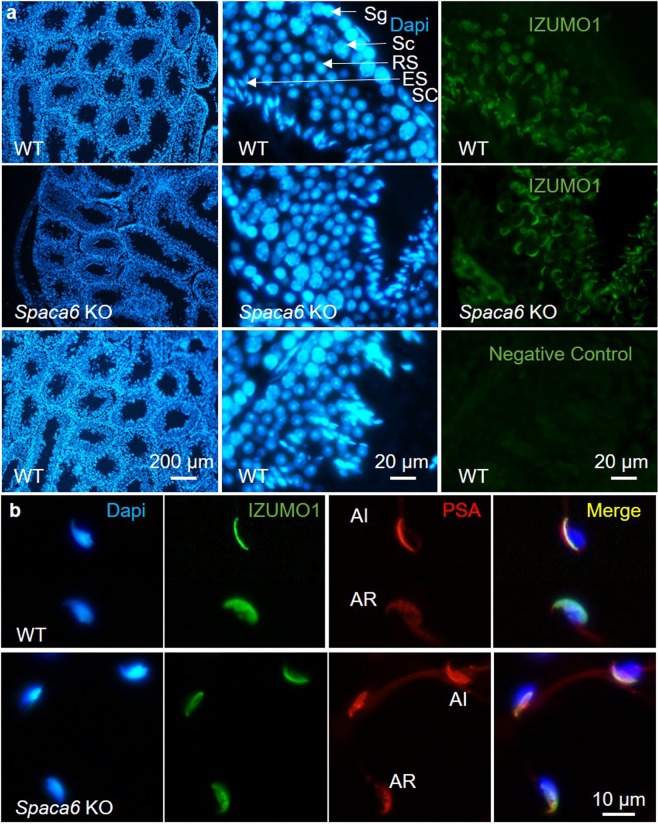
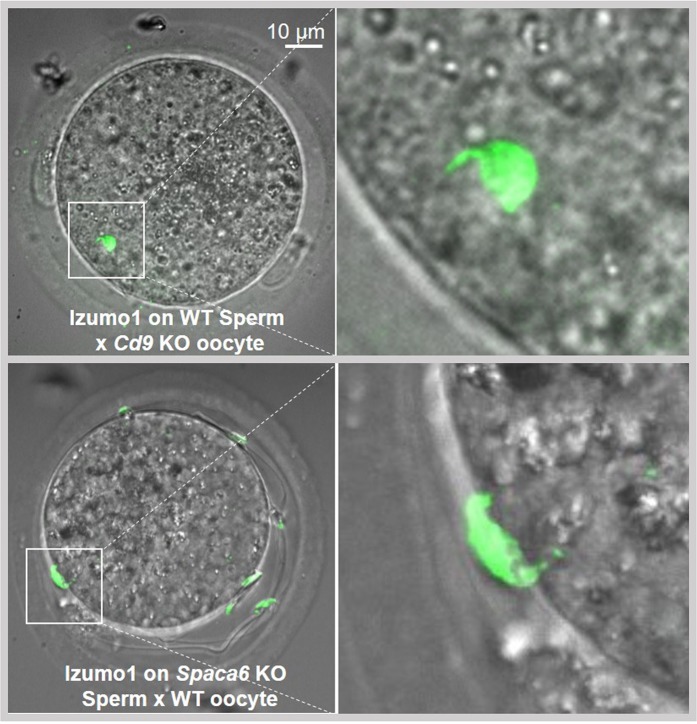
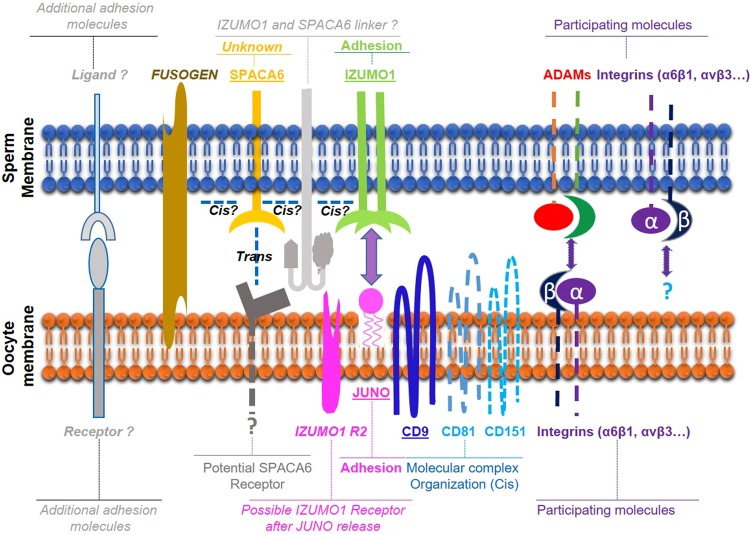
Three genes are known to be essential for gamete adhesion/fusion (Cd9, Izumo1 and Juno). Here, we confirmed that Spaca6 null males are infertile and showed that their sperm accumulate in the perivitelline space but are unable to fuse with oocyte. Like IZUMO1, SPACA6 which is expressed by human sperm, is remained on the equatorial segment after acrosomal reaction and is involved in human fertilization since an anti-SPACA6 antibody inhibited it. Despite the similarity of the phenotypes caused by Spaca6 and Izumo1 knockouts, these are not redundant and the essential relocation of IZUMO1 is not affected by the lack of SPACA6. We propose a model in which IZUMO1 and SPACA6 would be part of a molecular complex necessary for gamete fusion and that their concomitant presence would be required for the recruitment of another essential molecular actor, such as a fusogen, for the fusion to take place.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
