

LXR Activation Induces a Proinflammatory Trained Innate Immunity-Phenotype in Human Monocytes
- PMID: 32210962
- PMCID: PMC7077358
- DOI: 10.3389/fimmu.2020.00353
LXR Activation Induces a Proinflammatory Trained Innate Immunity-Phenotype in Human Monocytes
Abstract
Objectives: The concept of trained innate immunity describes a long-term proinflammatory memory in innate immune cells. Trained innate immunity is regulated through reprogramming of cellular metabolic pathways including cholesterol and fatty acid synthesis. Here, we have analyzed the role of Liver X Receptor (LXR), a key regulator of cholesterol and fatty acid homeostasis, in trained innate immunity.
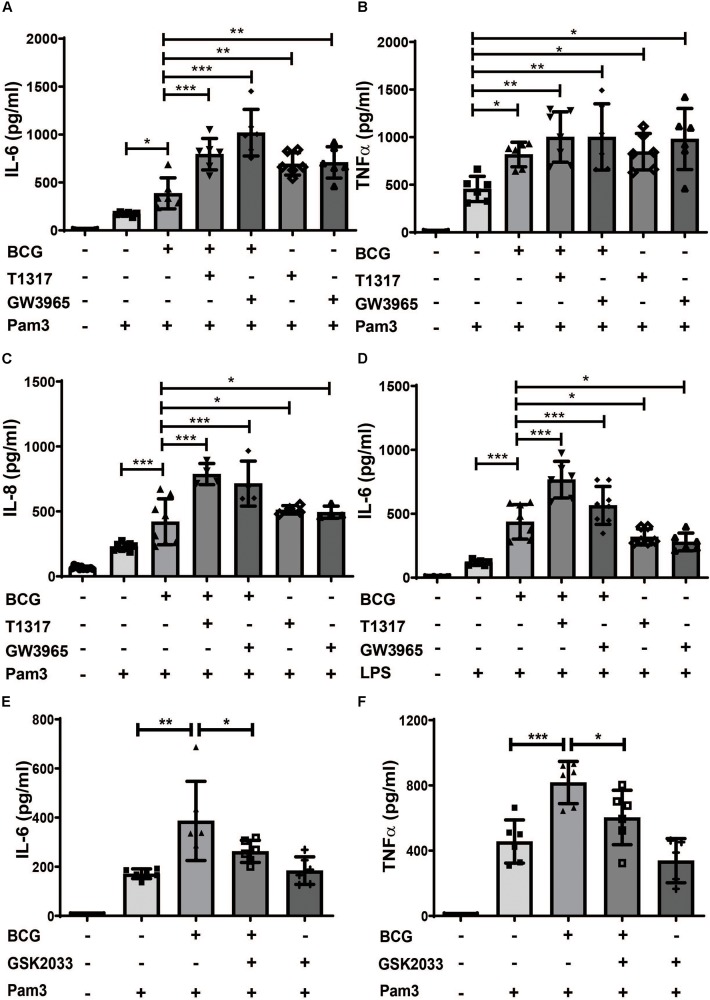
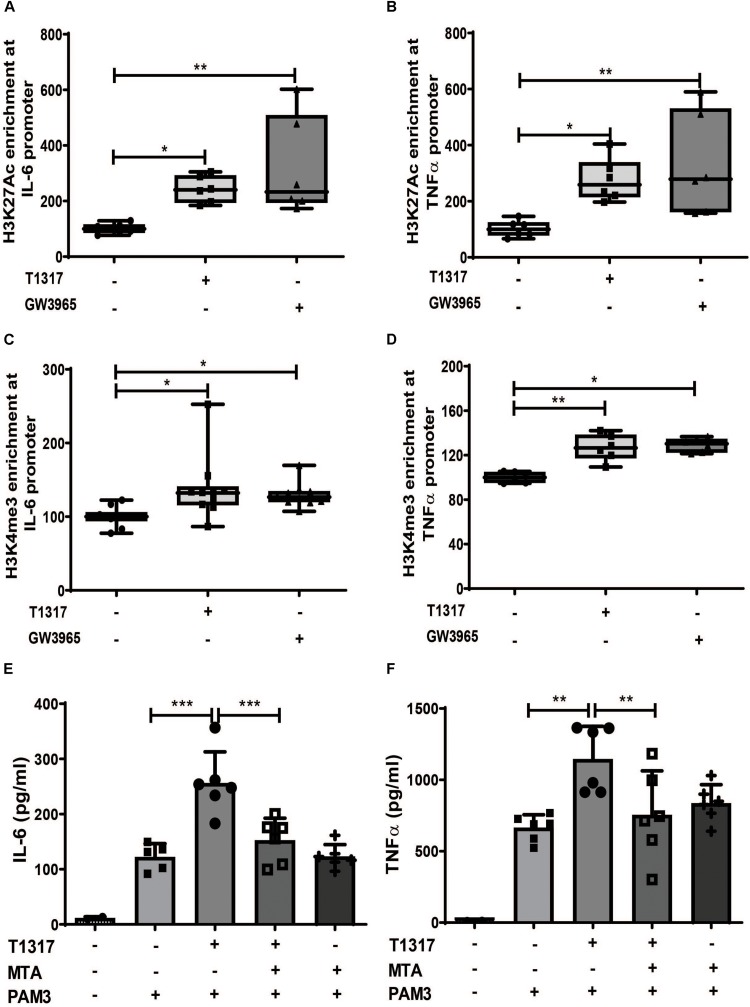
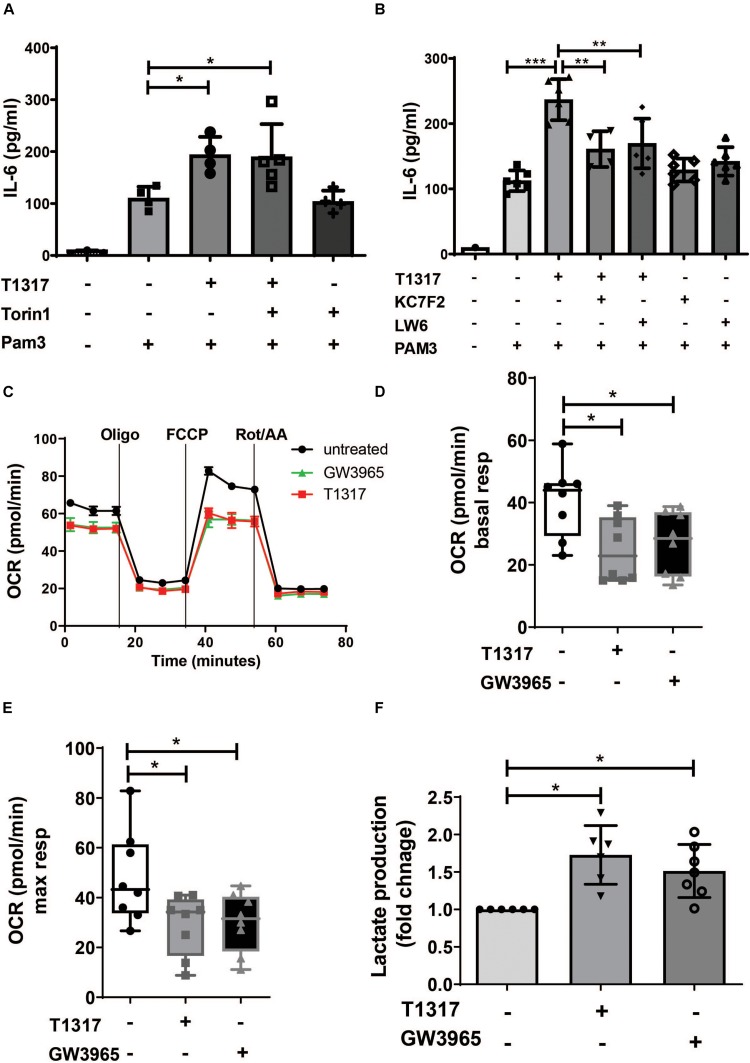
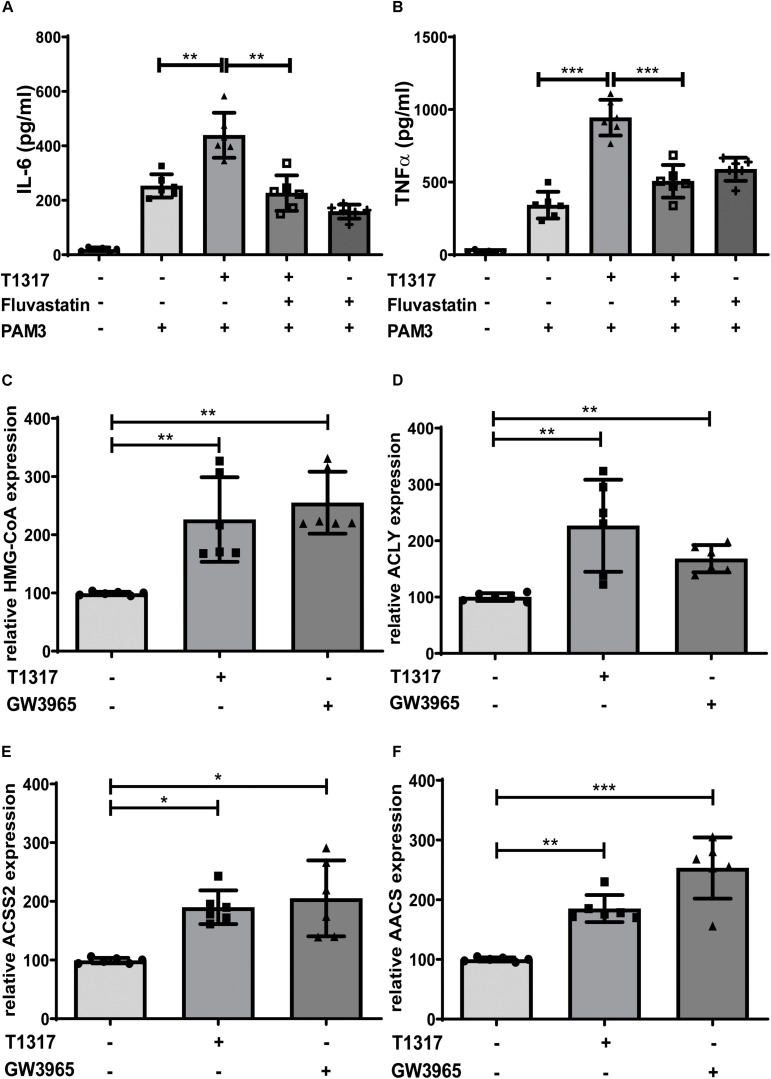
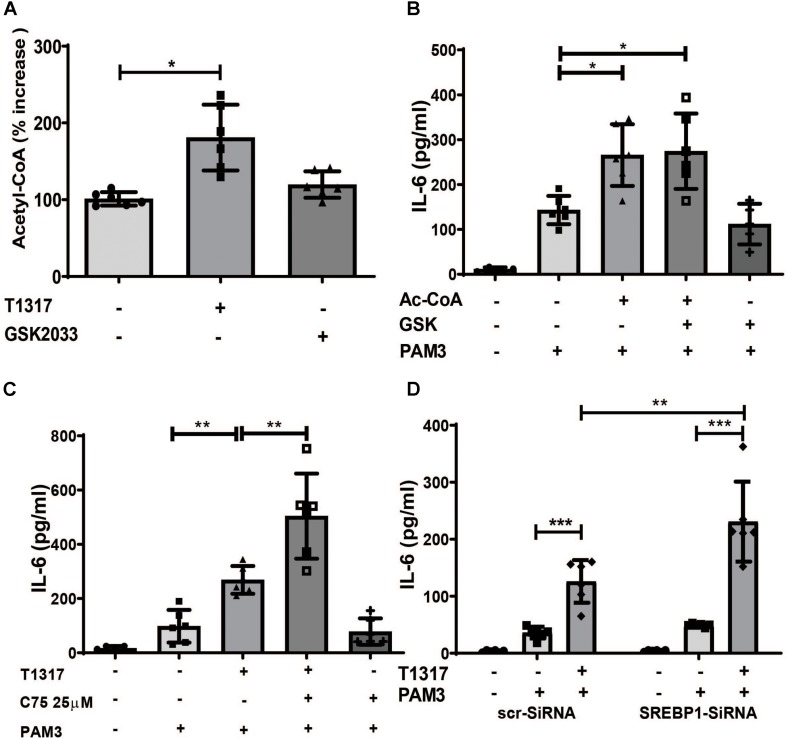
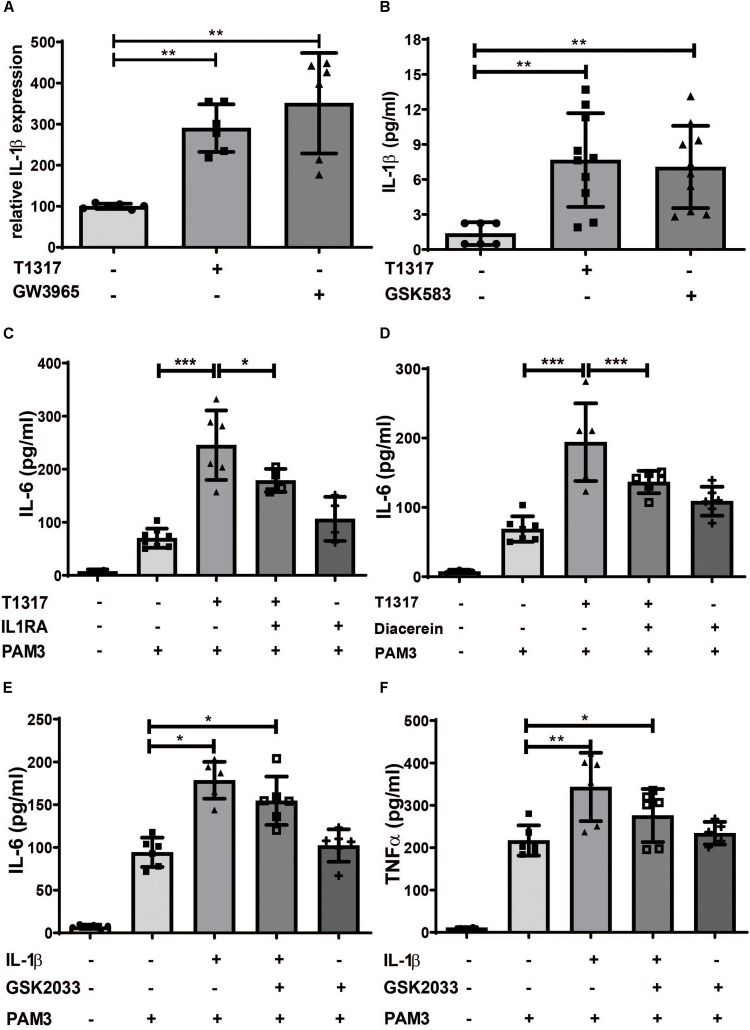
Methods and results: Human monocytes were isolated and incubated with different stimuli for 24 h, including LXR agonists, antagonists and Bacillus Calmette-Guerin (BCG) vaccine. After 5 days resting time, cells were restimulated with the TLR2-agonist Pam3cys. LXR activation did not only increase BCG trained immunity, but also induced a long-term inflammatory activation by itself. This inflammatory activation by LXR agonists was accompanied by characteristic features of trained innate immunity, such as activating histone marks on inflammatory gene promoters and metabolic reprogramming with increased lactate production and decreased oxygen consumption rate. Mechanistically, LXR priming increased cellular acetyl-CoA levels and was dependent on the activation of the mevalonate pathway and IL-1β signaling. In contrast to mevalonate pathway inhibition, blocking fatty acid synthesis further increased proinflammatory priming by LXR.
Conclusion: We demonstrate that LXR activation induces a proinflammatory trained immunity phenotype in human monocytes through epigenetic and metabolic reprogramming. Our data reveal important novel aspects of LXR signaling in innate immunity.
Keywords: LXR; acetyl-CoA; inflammation; monocytes; trained innate immunity.
Copyright © 2020 Sohrabi, Sonntag, Braun, Lagache, Liebmann, Klotz, Godfrey, Kahles, Waltenberger and Findeisen.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources