

HER2-Mediated Internalization of Cytotoxic Agents in ERBB2 Amplified or Mutant Lung Cancers
- PMID: 32213539
- PMCID: PMC7196485
- DOI: 10.1158/2159-8290.CD-20-0215
HER2-Mediated Internalization of Cytotoxic Agents in ERBB2 Amplified or Mutant Lung Cancers
Abstract
Amplification of and oncogenic mutations in ERBB2, the gene encoding the HER2 receptor tyrosine kinase, promote receptor hyperactivation and tumor growth. Here we demonstrate that HER2 ubiquitination and internalization, rather than its overexpression, are key mechanisms underlying endocytosis and consequent efficacy of the anti-HER2 antibody-drug conjugates (ADC) ado-trastuzumab emtansine (T-DM1) and trastuzumab deruxtecan (T-DXd) in lung cancer cell lines and patient-derived xenograft models. These data translated into a 51% response rate in a clinical trial of T-DM1 in 49 patients with ERBB2-amplified or -mutant lung cancers. We show that cotreatment with irreversible pan-HER inhibitors enhances receptor ubiquitination and consequent ADC internalization and efficacy. We also demonstrate that ADC switching to T-DXd, which harbors a different cytotoxic payload, achieves durable responses in a patient with lung cancer and corresponding xenograft model developing resistance to T-DM1. Our findings may help guide future clinical trials and expand the field of ADC as cancer therapy. SIGNIFICANCE: T-DM1 is clinically effective in lung cancers with amplification of or mutations in ERBB2. This activity is enhanced by cotreatment with irreversible pan-HER inhibitors, or ADC switching to T-DXd. These results may help address unmet needs of patients with HER2-activated tumors and no approved targeted therapy.See related commentary by Rolfo and Russo, p. 643.This article is highlighted in the In This Issue feature, p. 627.
©2020 American Association for Cancer Research.
Conflict of interest statement
Competing interests
M.S. has received research funds from AstraZeneca, Puma Biotechnology, Daiichi Sankyo, Immunomedics, Targimmune and Menarini Ricerche. He is in the scientific advisory board (SAB) of Menarini Ricerche and Bioscience Institute and is a cofounder of Medendi.org. D.B.S. has served as consulted/received honoraria from Pfizer, Loxo Oncology, Illumina, Lilly Oncology and Vivideon Therapeutics. S.M. consulted for Boehringer-Ingelheim. M.S. and B.T.L. have two pending institutional patents at Memorial Sloan Kettering Cancer Center (US62/685,057, US62/514,661). B.T.L. has served as a consultant/advisory board member for Roche/Genentech, Biosceptre International, Thermo Fisher Scientific, Mersana Therapeutics, Hengrui Therapeutics, Guardant Health and has received research funds to his institution from Roche/Genentech, Daiichi Sankyo, Hengrui Therapeutics, Illumina, Guardant Health, BioMed Valley Discoveries, AstraZeneca, GRAIL, MORE Health, Amgen and Lilly. C.M.R. is a consultant/advisory board member for AbbVie, Amgen, Ascentage, AstraZeneca, Bristol-Myers Squibb, Celgene, Daiichi Sankyo, Genentech/Roche, Ipsen, Loxo, PharmaMar, Bridge Medicine, and Harpoon. R.A.F. receives research funds from Puma Biotechnology and Eisai Pharmaceuticals. N.U.L. has received research support (to institution) from Pfizer, Genentech, Seattle Genetics, and Merck, and has been a consultant for Puma, Daichii, and Seattle Genetics. D.M.H. reports receiving commercial research grants from Loxo, PUMA Biotechnology, AstraZeneca, and Bayer Pharmaceuticals, and is a consultant/advisory board member for Chugai Pharma, CytomX Therapeutics, Boehringer Ingelheim, AstraZeneca, Pfizer, Bayer Pharmaceuticals, and Genentech/Roche. G.A.U. is a consultant for Sanofi and receives research support for Sanofi, Novartis, Genentech, and Puma Biotechnology. M.G.K. has received consulting fees from AstraZeneca, Pfizer, and Regeneron. He has received research funding from The National Cancer Institute (USA), The Lung Cancer Research Foundation, Genentech Roche, and PUMA Biotechnology and honoraria He has received honoraria for participation in educational programs from WebMD, OncLive, Physicians Education Resources, Prime Oncology, Intellisphere, Creative Educational Concepts, Peerview, i3 Health, Paradigm Medical Communications, AXIS, Carvive Systems, AstraZeneca, and Research to Practice. N.R. is on the SAB and receives research funding from Chugai, on the SAB and owns equity in Beigene, and Fortress. N.R. is also on the SAB of Daiichi-Sankyo, Astra-Zeneca-MedImmune, and F-Prime, and is a past SAB member of Millenium-Takeda, Kadmon, Kura, and Araxes. N.R. is a consultant to Novartis, Boehringer Ingelheim, Tarveda, and Foresight and consulted in the last three years with Eli Lilly, Merrimack, Kura Oncology, Araxes, and Kadman. N.R. owns equity in ZaiLab, Kura Oncology, Araxes, and Kadman. N.R. also collaborates with Plexxikon. M.F.B. declares consulting/advisory board activities for Roche. J.S.R.-F. is a consultant of Goldman Sachs Merchant Bank and REPARE Therapeutics, a member of the SAB of Volition RX and Paige.AI, and an ad hoc member of the SABs of Roche Tissue Diagnostics, Ventana, InVicro, Novartis and Genentech. D.R.J. is a senior consultant for Diffusions Pharmaceuticals, Consultant for AstraZeneca, and Merck. M.O. is a consultant/advisory board member for PharmaMar, Novartis and Targeted Oncology. He received research funds from Bristol-Myers Squibb (Inst) and Merck Sharp & Dohme (Inst). J.M.I. owns equity in LumaCyte, LLC. F.C. is an AstraZeneca employee. S.T., W-L.L. and A.B. are mProbe Inc. employees.
Memorial Sloan Kettering has an institutional agreement with IBM for Watson For Oncology and receives royalties from IBM.
No potential conflicts of interests were disclosed by the other authors.
Figures
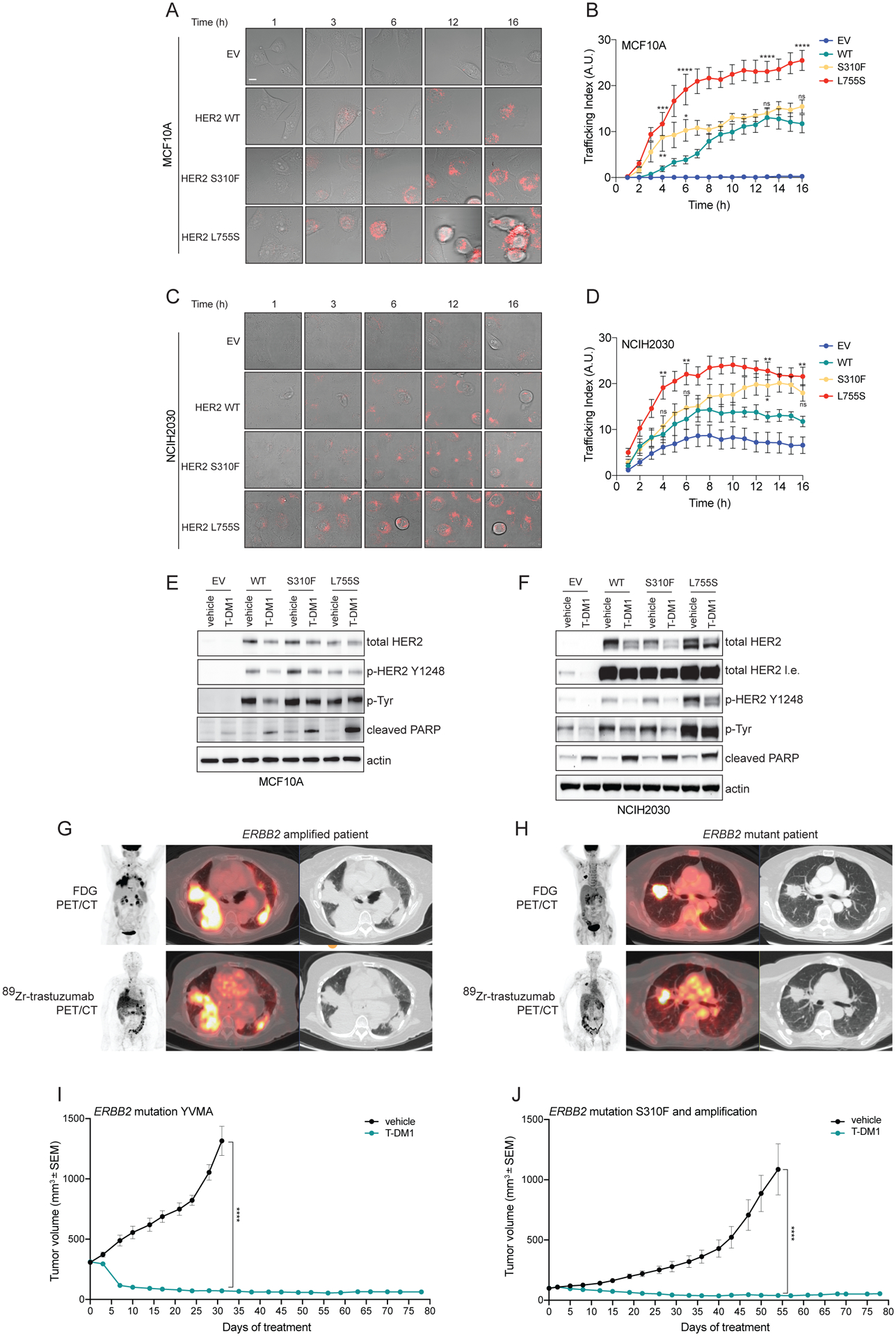
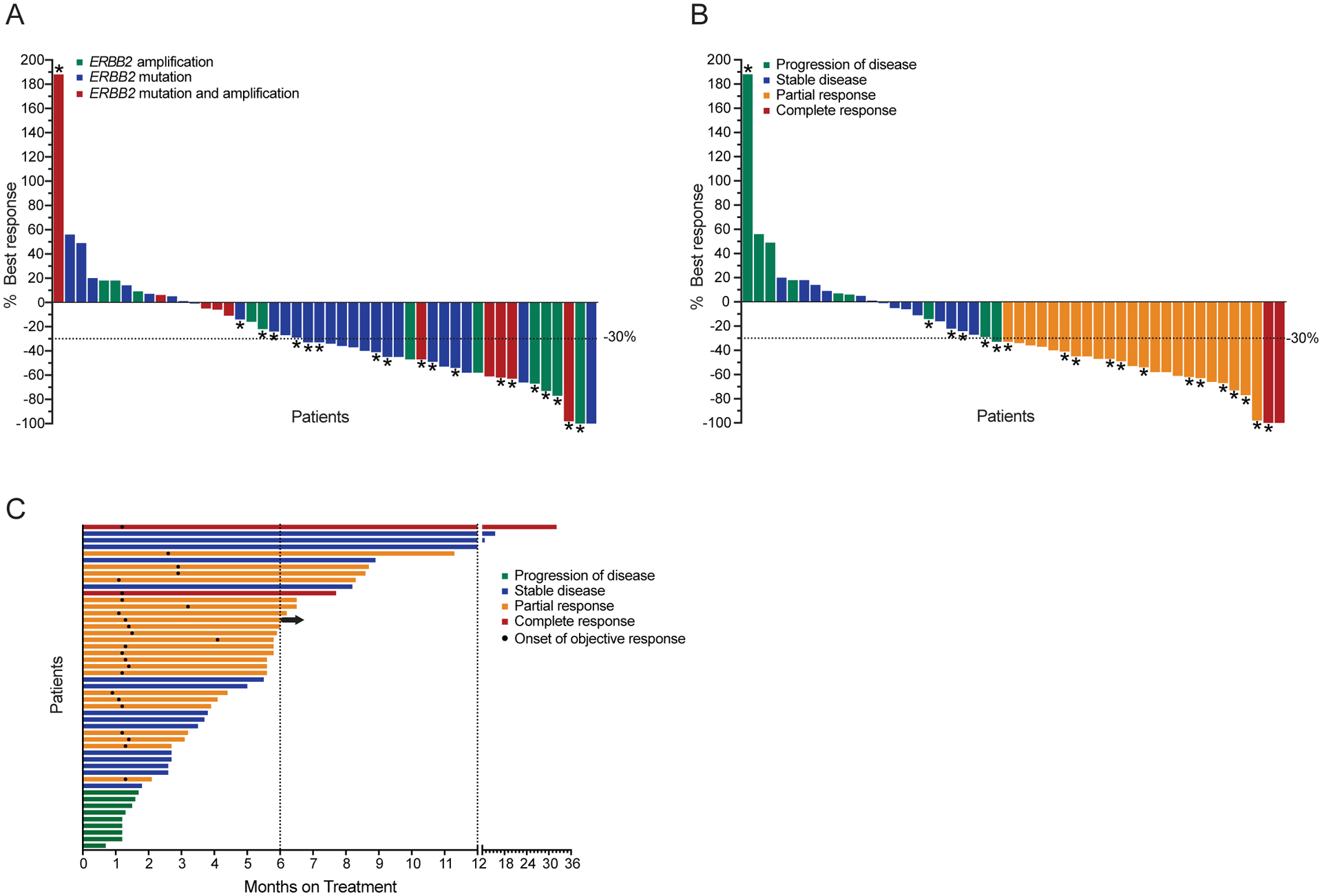
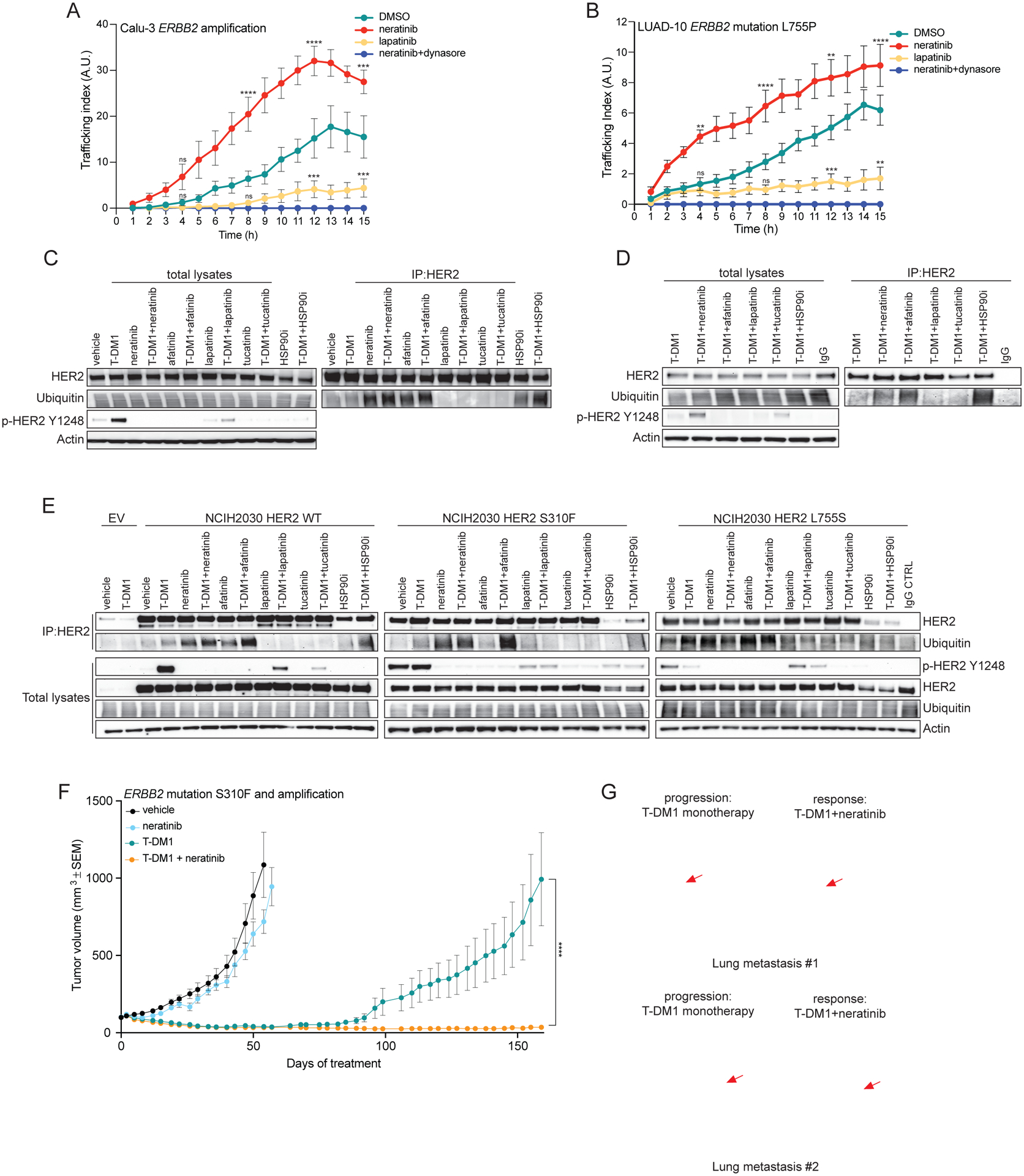
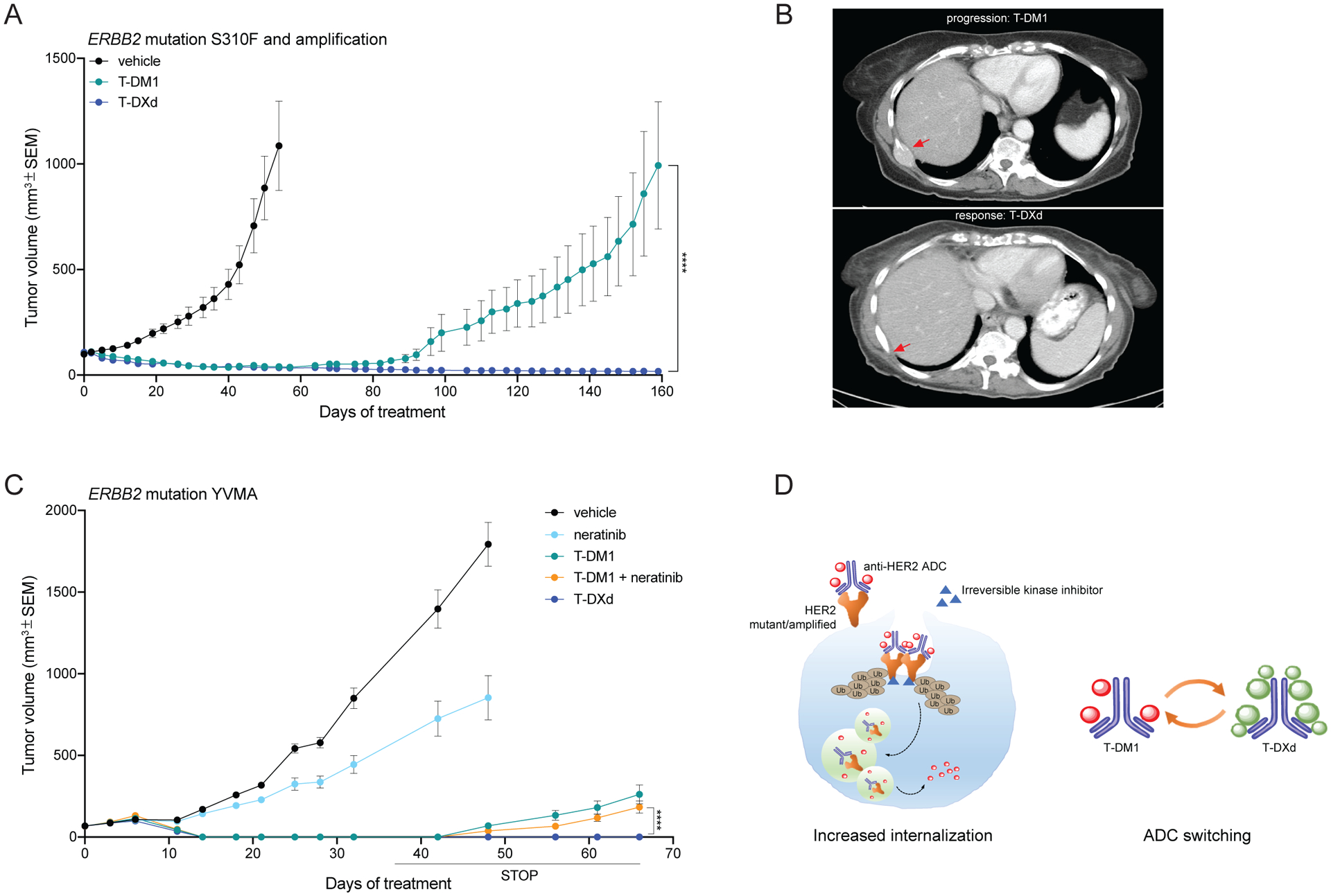
Comment in
-
HER2 Mutations in Non-Small Cell Lung Cancer: A Herculean Effort to Hit the Target.Cancer Discov. 2020 May;10(5):643-645. doi: 10.1158/2159-8290.CD-20-0225. Cancer Discov. 2020. PMID: 32357968
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
