

Metastasis of cholangiocarcinoma is promoted by extended high-mannose glycans
- PMID: 32213588
- PMCID: PMC7148571
- DOI: 10.1073/pnas.1916498117
Metastasis of cholangiocarcinoma is promoted by extended high-mannose glycans
Abstract
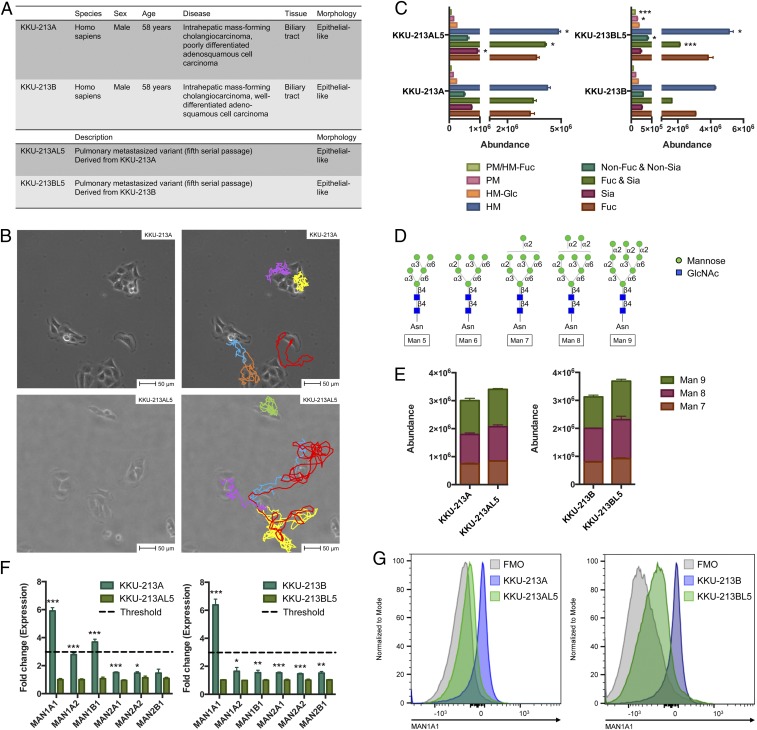
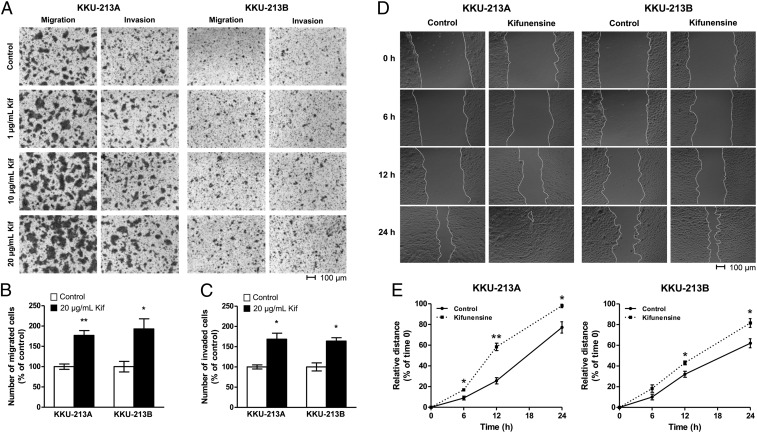
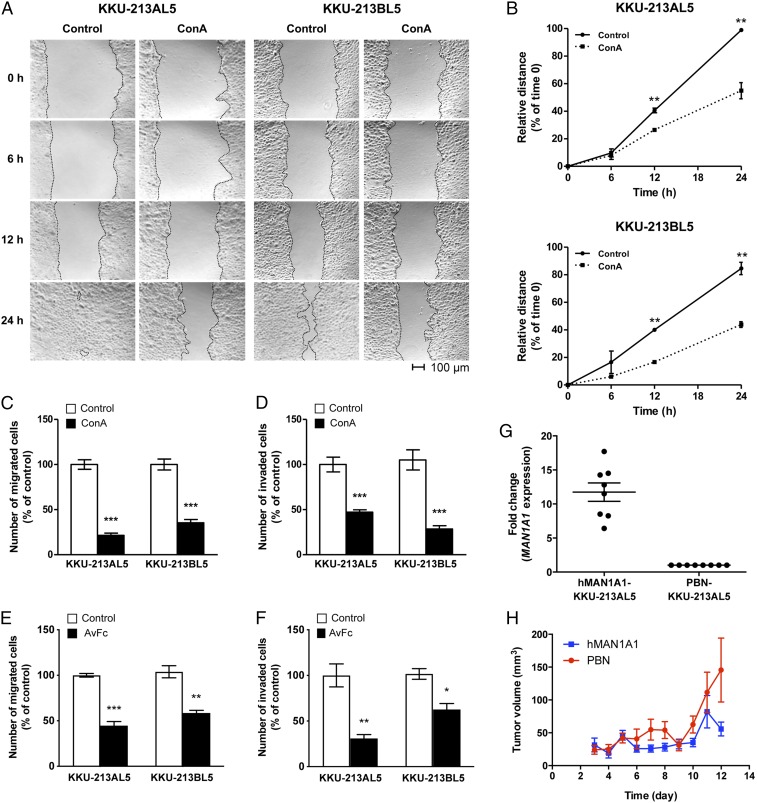
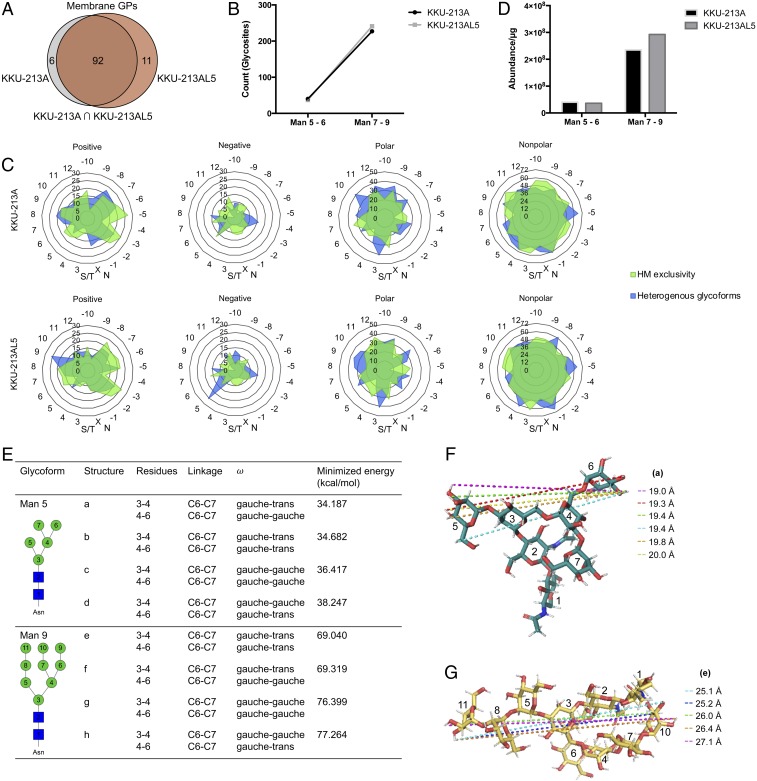
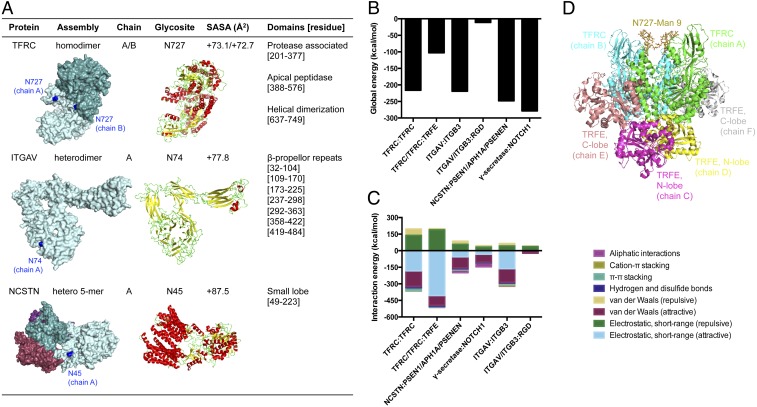
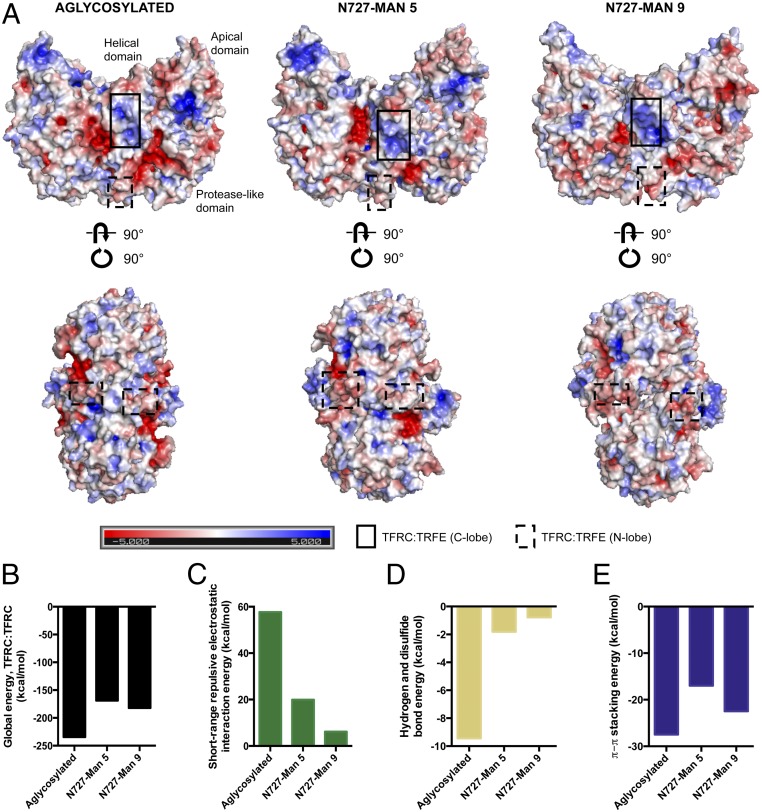
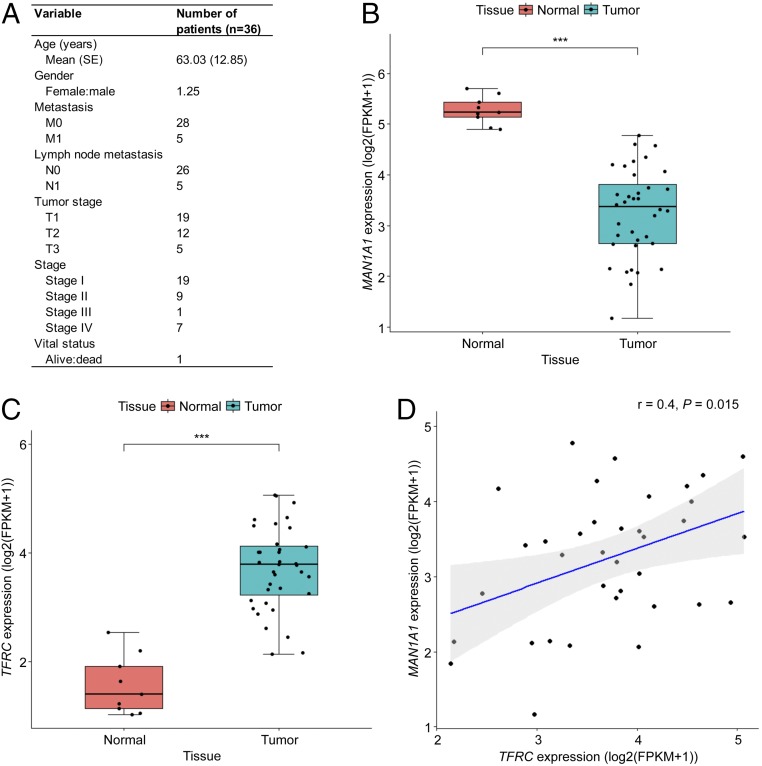
Membrane-bound oligosaccharides form the interfacial boundary between the cell and its environment, mediating processes such as adhesion and signaling. These structures can undergo dynamic changes in composition and expression based on cell type, external stimuli, and genetic factors. Glycosylation, therefore, is a promising target of therapeutic interventions for presently incurable forms of advanced cancer. Here, we show that cholangiocarcinoma metastasis is characterized by down-regulation of the Golgi α-mannosidase I coding gene MAN1A1, leading to elevation of extended high-mannose glycans with terminating α-1,2-mannose residues. Subsequent reshaping of the glycome by inhibiting α-mannosidase I resulted in significantly higher migratory and invasive capabilities while masking cell surface mannosylation suppressed metastasis-related phenotypes. Exclusive elucidation of differentially expressed membrane glycoproteins and molecular modeling suggested that extended high-mannose glycosylation at the helical domain of transferrin receptor protein 1 promotes conformational changes that improve noncovalent interaction energies and lead to enhancement of cell migration in metastatic cholangiocarcinoma. The results provide support that α-1,2-mannosylated N-glycans present on cancer cell membrane proteins may serve as therapeutic targets for preventing metastasis.
Keywords: cholangiocarcinoma; glycosylation; mass spectrometry; membrane proteins; metastasis.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Bogenrieder T., Herlyn M., Axis of evil: Molecular mechanisms of cancer metastasis. Oncogene 22, 6524–6536 (2003). - PubMed
-
- Friedl P., Wolf K., Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 3, 362–374 (2003). - PubMed
-
- Geiger T. R., Peeper D. S., Metastasis mechanisms. Biochim. Biophys. Acta 1796, 293–308 (2009). - PubMed
-
- Khan S. A., Thomas H. C., Davidson B. R., Taylor-Robinson S. D., Cholangiocarcinoma. Lancet 366, 1303–1314 (2005). - PubMed
-
- Dube D. H., Bertozzi C. R., Glycans in cancer and inflammation'–potential for therapeutics and diagnostics. Nat. Rev. Drug Discov. 4, 477–488 (2005). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
