

Fluorescence-Detected Conformational Changes in Duplex DNA in Open Complex Formation by Escherichia coli RNA Polymerase: Upstream Wrapping and Downstream Bending Precede Clamp Opening and Insertion of the Downstream Duplex
- PMID: 32216369
- PMCID: PMC7269339
- DOI: 10.1021/acs.biochem.0c00098
Fluorescence-Detected Conformational Changes in Duplex DNA in Open Complex Formation by Escherichia coli RNA Polymerase: Upstream Wrapping and Downstream Bending Precede Clamp Opening and Insertion of the Downstream Duplex
Abstract
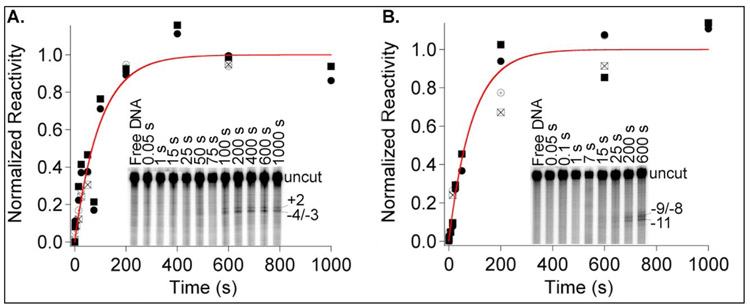
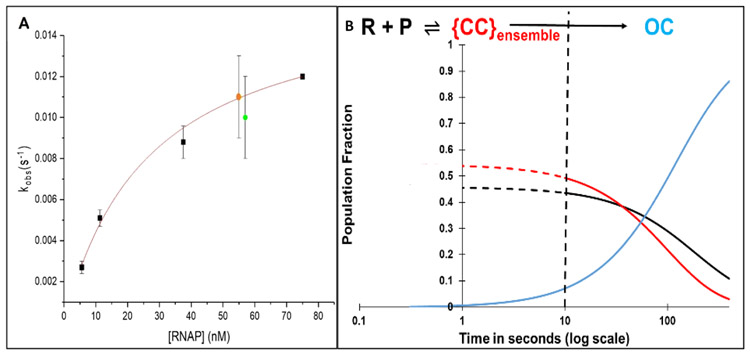
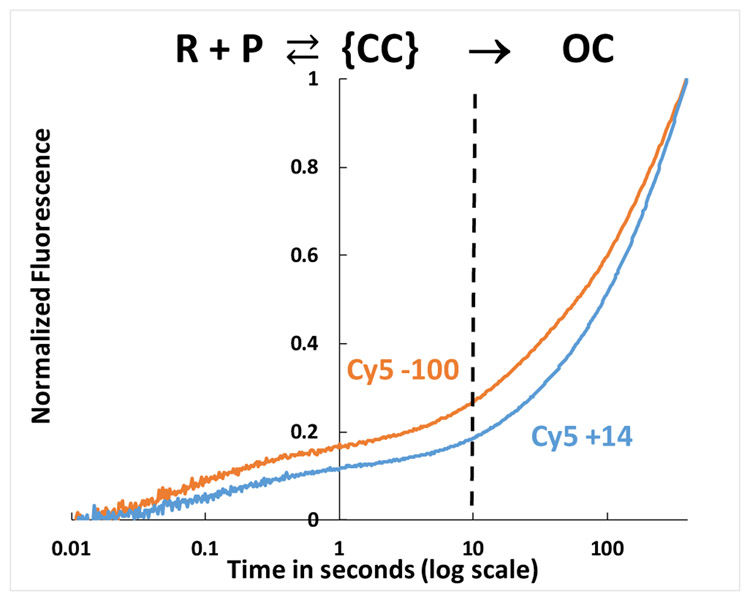
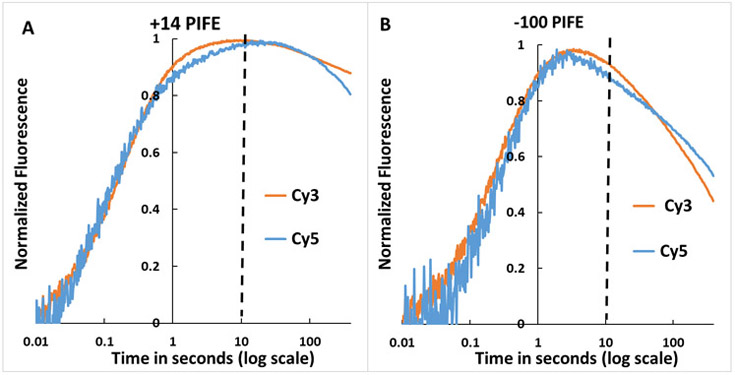
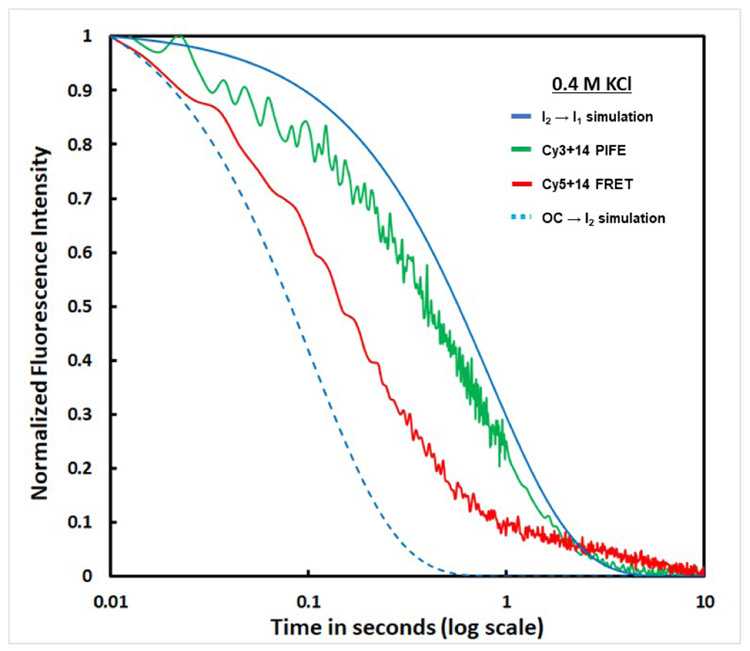
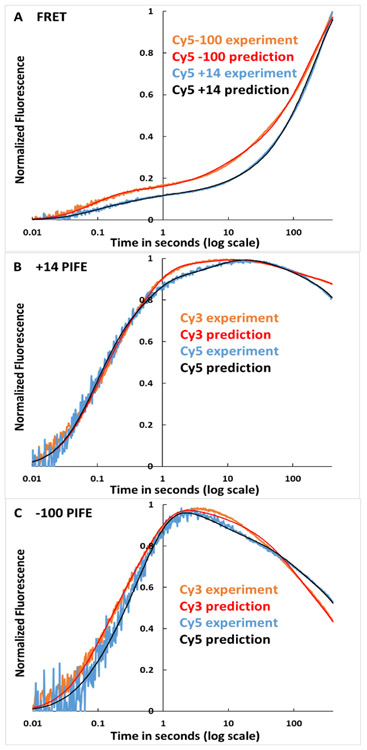
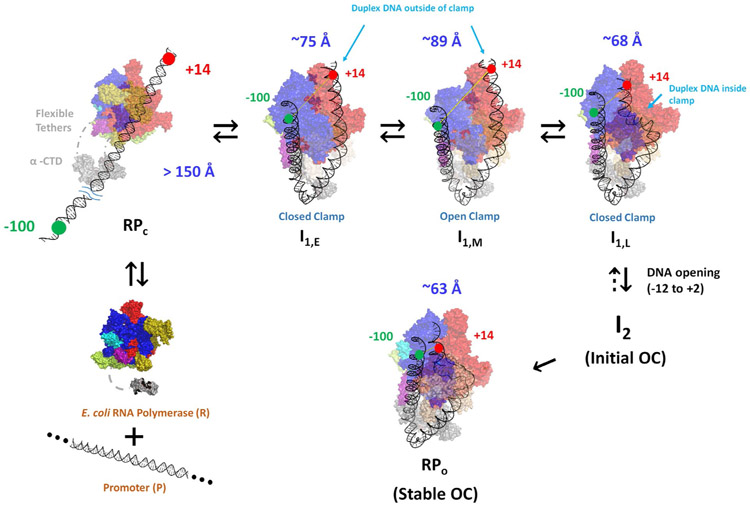
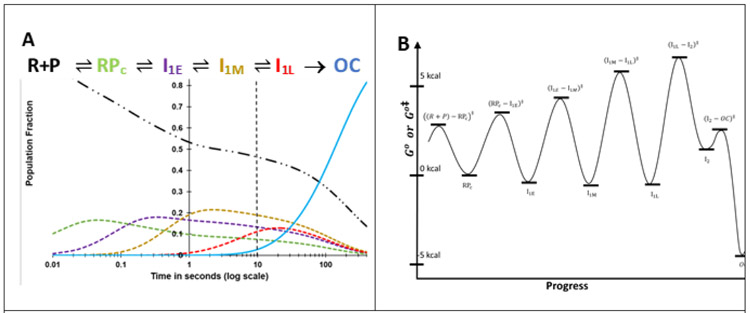
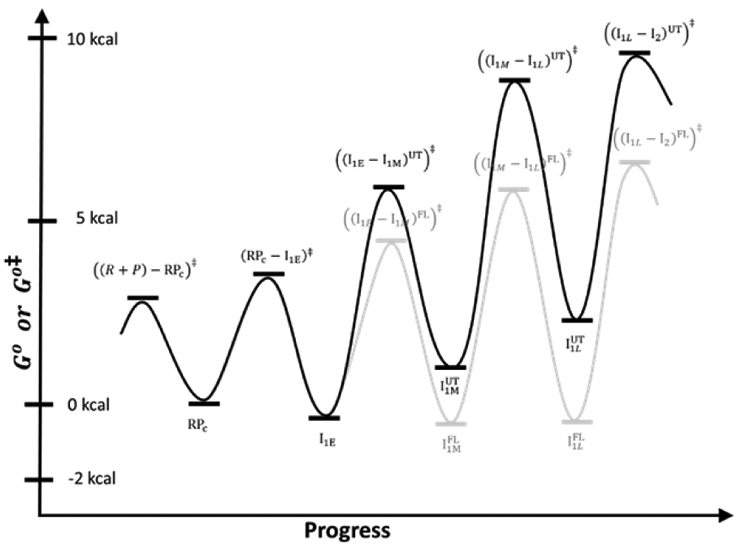
FRET (fluorescence resonance energy transfer) between far-upstream (-100) and downstream (+14) cyanine dyes (Cy3, Cy5) showed extensive bending and wrapping of λPR promoter DNA on Escherichia coli RNA polymerase (RNAP) in closed and open complexes (CC and OC, respectively). Here we determine the kinetics and mechanism of DNA bending and wrapping by FRET and of formation of RNAP contacts with -100 and +14 DNA by single-dye protein-induced fluorescence enhancement (PIFE). FRET and PIFE kinetics exhibit two phases: rapidly reversible steps forming a CC ensemble ({CC}) of four intermediates [initial (RPC), early (I1E), mid (I1M), and late (I1L)], followed by conversion of {CC} to OC via I1L. FRET and PIFE are first observed for I1E, not RPc. FRET and PIFE together reveal large-scale bending and wrapping of upstream and downstream DNA as RPC advances to I1E, decreasing the Cy3-Cy5 distance to ∼75 Å and making RNAP-DNA contacts at -100 and +14. We propose that far-upstream DNA wraps on the upper β'-clamp while downstream DNA contacts the top of the β-pincer in I1E. Converting I1E to I1M (∼1 s time scale) reduces FRET efficiency with little change in -100 or +14 PIFE, interpreted as clamp opening that moves far-upstream DNA (on β') away from downstream DNA (on β) to increase the Cy3-Cy5 distance by ∼14 Å. FRET increases greatly in converting I1M to I1L, indicating bending of downstream duplex DNA into the clamp and clamp closing to reduce the Cy3-Cy5 distance by ∼21 Å. In the subsequent rate-determining DNA-opening step, in which the clamp may also open, I1L is converted to the initial unstable OC (I2). Implications for facilitation of CC-to-OC isomerization by upstream DNA and upstream binding, DNA-bending transcription activators are discussed.
Figures
References
-
- McClure WR (1985) Mechanism and control of transcription initiation in prokaryotes, Annual Review of Biochemistry 54, 171–204. - PubMed
-
- Record MT Jr., Reznikoff WS, Craig ML, Mcquade KL and Schlax PJ. (1996) Escherichia coli RNA Polymerase (E sigma70), Promoters, and the Kinetics of the Steps of Transcription Initiation., Second Edition of Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology, 792–821.
-
- Saecker RM, Tsodikov OV, Mcquade KL, Schlax PE Jr, Capp MW, and Record MT Jr. (2002) Kinetic studies and structural models of the association of E. coli σ70 RNA polymerase with the λPR promoter: large scale conformational changes in forming the kinetically significant intermediates, Journal of Molecular Biology 319, 649–671. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
