

Development of a new oral poliovirus vaccine for the eradication end game using codon deoptimization
- PMID: 32218998
- PMCID: PMC7083942
- DOI: 10.1038/s41541-020-0176-7
Development of a new oral poliovirus vaccine for the eradication end game using codon deoptimization
Abstract
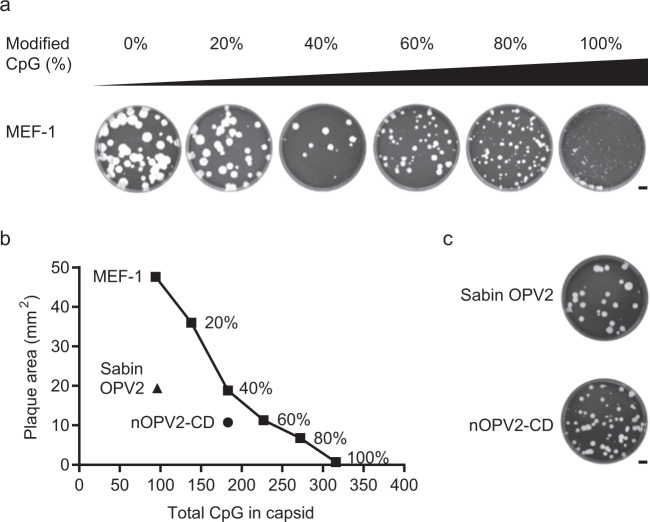
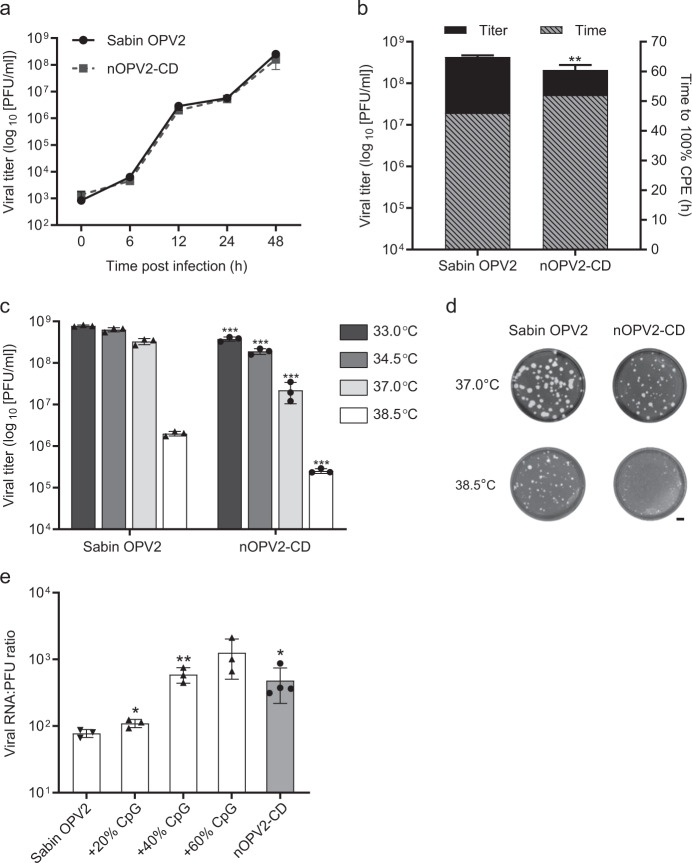
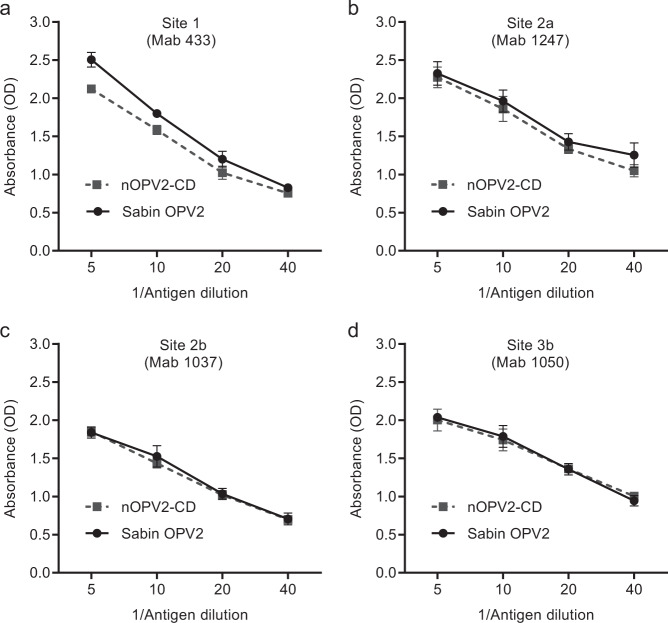
Enormous progress has been made in global efforts to eradicate poliovirus, using live-attenuated Sabin oral poliovirus vaccine (OPV). However, as the incidence of disease due to wild poliovirus has declined, vaccine-derived poliovirus (VDPV) has emerged in areas of low-vaccine coverage. Coordinated global cessation of routine, type 2 Sabin OPV (OPV2) use has not resulted in fewer VDPV outbreaks, and continued OPV use in outbreak-response campaigns has seeded new emergences in low-coverage areas. The limitations of existing vaccines and current eradication challenges warranted development of more genetically stable OPV strains, most urgently for OPV2. Here, we report using codon deoptimization to further attenuate Sabin OPV2 by changing preferred codons across the capsid to non-preferred, synonymous codons. Additional modifications to the 5' untranslated region stabilized known virulence determinants. Testing of this codon-deoptimized new OPV2 candidate (nOPV2-CD) in cell and animal models demonstrated that nOPV2-CD is highly attenuated, grows sufficiently for vaccine manufacture, is antigenically indistinguishable from Sabin OPV2, induces neutralizing antibodies as effectively as Sabin OPV2, and unlike Sabin OPV2 is genetically stable and maintains an attenuation phenotype. In-human clinical trials of nOPV2-CD are ongoing, with potential for nOPV strains to serve as critical vaccine tools for achieving and maintaining polio eradication.
Keywords: Live attenuated vaccines; Policy and public health in microbiology; Vaccines; Virology.
© This is a U.S. government work and not under copyright protection in the U.S.; foreign copyright protection may apply 2020.
Conflict of interest statement
Competing interestsR.C., J.S., O.K., and C.C.B are listed as co-inventors on a US patent (“Modulation of replicative fitness by deoptimization of synonymous codons,” US patent #8,846,051) held by the US Government. Similar patents have been issued in Europe, Hong Kong, India, Canada, and Australia. The remaining authors declare no competing interests.
Figures
References
-
- Kew O, Pallansch M. Breaking the last chains of poliovirus transmission: progress and challenges in global polio eradication. Annu. Rev. Virol. 2018;5:427–451. - PubMed
-
- Dunn G, Begg NT, Cammack N, Minor PD. Virus excretion and mutation by infants following primary vaccination with live oral poliovaccine from two sources. J. Med. Virol. 1990;32:92–95. - PubMed
-
- Alexander LN, et al. Vaccine policy changes and epidemiology of poliomyelitis in the United States. J. Am. Med. Assoc. 2004;292:1696–1701. - PubMed
