

Synergistic Cues from Diverse Bacteria Enhance Multicellular Development in a Choanoflagellate
- PMID: 32220848
- PMCID: PMC7237790
- DOI: 10.1128/AEM.02920-19
Synergistic Cues from Diverse Bacteria Enhance Multicellular Development in a Choanoflagellate
Abstract
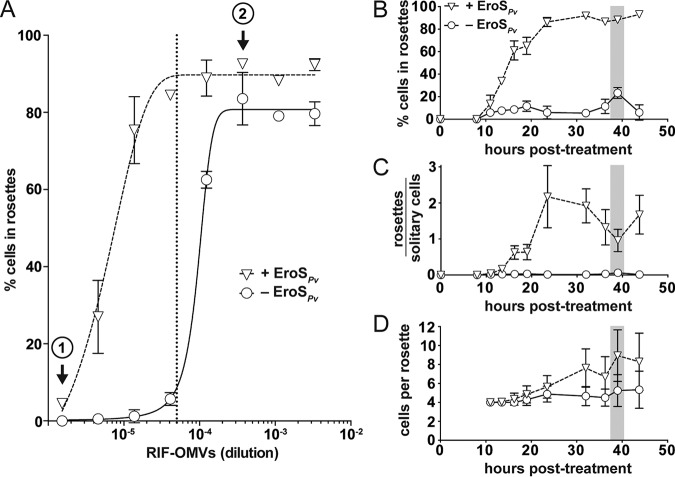
Bacteria regulate the life histories of diverse eukaryotes, but relatively little is known about how eukaryotes interpret and respond to multiple bacterial cues encountered simultaneously. To explore how a eukaryote might respond to a combination of bioactive molecules from multiple bacteria, we treated the choanoflagellate Salpingoeca rosetta with two sets of bacterial cues, one that induces mating and another that induces multicellular development. We found that simultaneous exposure to both sets of cues enhanced multicellular development in S. rosetta, eliciting both larger multicellular colonies and an increase in the number of colonies. Thus, rather than conveying conflicting sets of information, these distinct bacterial cues synergize to augment multicellular development. This study demonstrates how a eukaryote can integrate and modulate its response to cues from diverse bacteria, underscoring the potential impact of complex microbial communities on eukaryotic life histories.IMPORTANCE Eukaryotic biology is profoundly influenced by interactions with diverse environmental and host-associated bacteria. However, it is not well understood how eukaryotes interpret multiple bacterial cues encountered simultaneously. This question has been challenging to address because of the complexity of many eukaryotic model systems and their associated bacterial communities. Here, we studied a close relative of animals, the choanoflagellate Salpingoeca rosetta, to explore how eukaryotes respond to diverse bacterial cues. We found that a bacterial chondroitinase that induces mating on its own can also synergize with bacterial lipids that induce multicellular "rosette" development. When encountered together, these cues enhance rosette development, resulting in both the formation of larger rosettes and an increase in the number of rosettes compared to rosette development in the absence of the chondroitinase. These findings highlight how synergistic interactions among bacterial cues can influence the biology of eukaryotes.
Keywords: EroS; RIF-1; Salpingoeca rosetta; choanoflagellate; chondroitinase; host microbe; multicellularity; outer membrane vesicles; rosette-inducing factor; sulfonolipid.
Copyright © 2020 Ireland et al.
Figures



References
-
- McFall-Ngai M, Hadfield MG, Bosch TCG, Carey HV, Domazet-Lošo T, Douglas AE, Dubilier N, Eberl G, Fukami T, Gilbert SF, Hentschel U, King N, Kjelleberg S, Knoll AH, Kremer N, Mazmanian SK, Metcalf JL, Nealson K, Pierce NE, Rawls JF, Reid A, Ruby EG, Rumpho M, Sanders JG, Tautz D, Wernegreen JJ. 2013. Animals in a bacterial world, a new imperative for the life sciences. Proc Natl Acad Sci U S A 110:3229–3236. doi: 10.1073/pnas.1218525110. - DOI - PMC - PubMed
Publication types
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
