

Systematic identification and elimination of flux bottlenecks in the aldehyde production pathway of Synechococcus elongatus PCC 7942
- PMID: 32222320
- PMCID: PMC7217728
- DOI: 10.1016/j.ymben.2020.03.007
Systematic identification and elimination of flux bottlenecks in the aldehyde production pathway of Synechococcus elongatus PCC 7942
Abstract
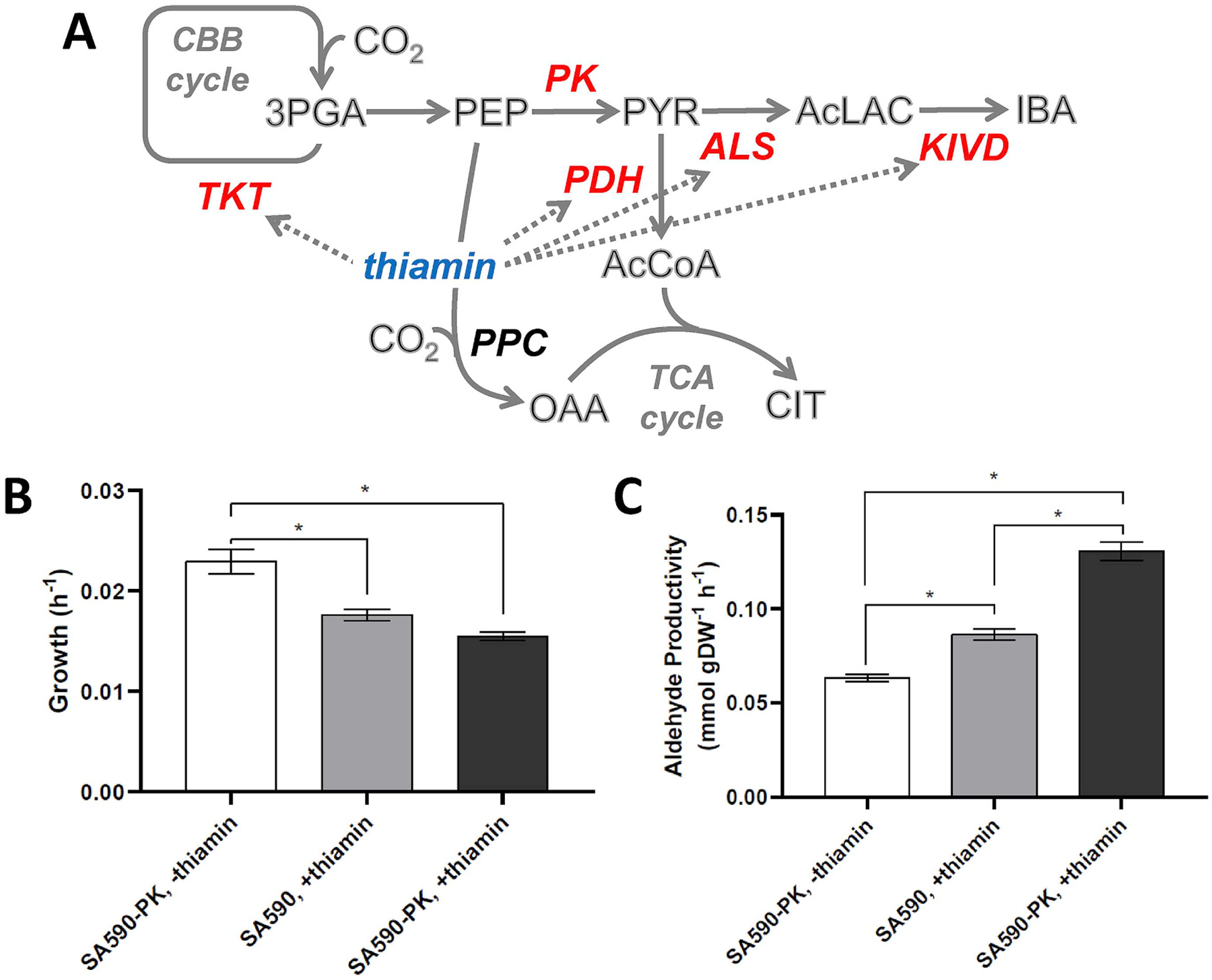
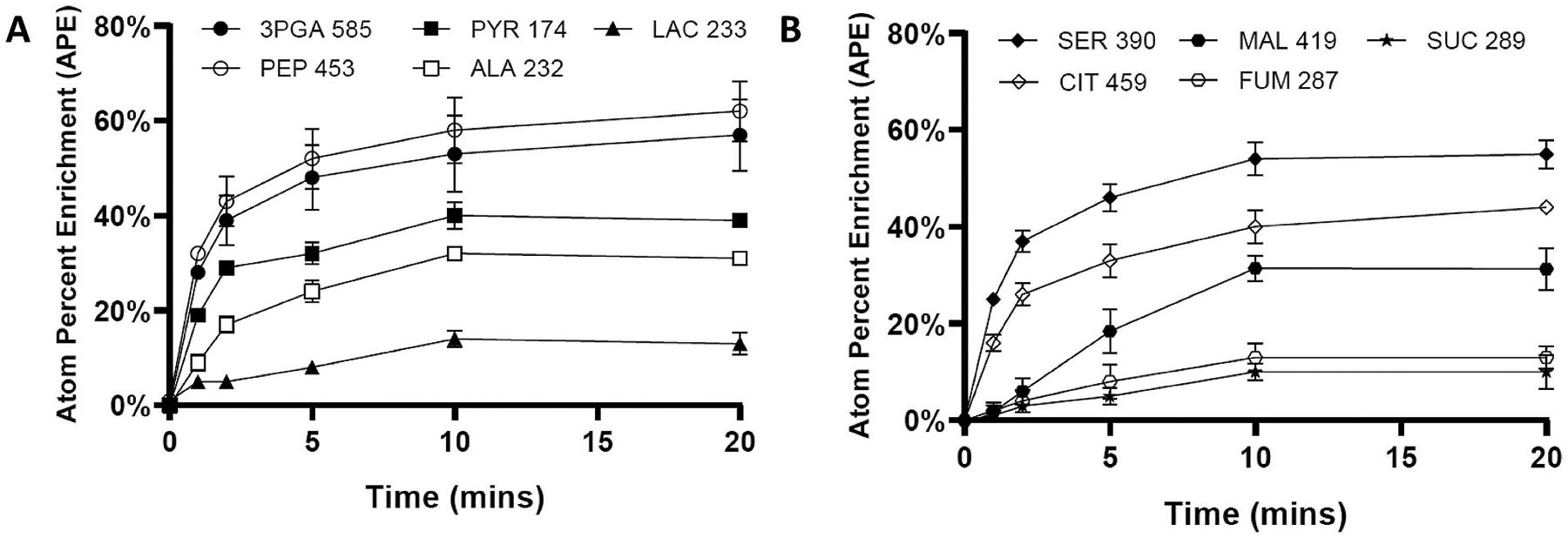
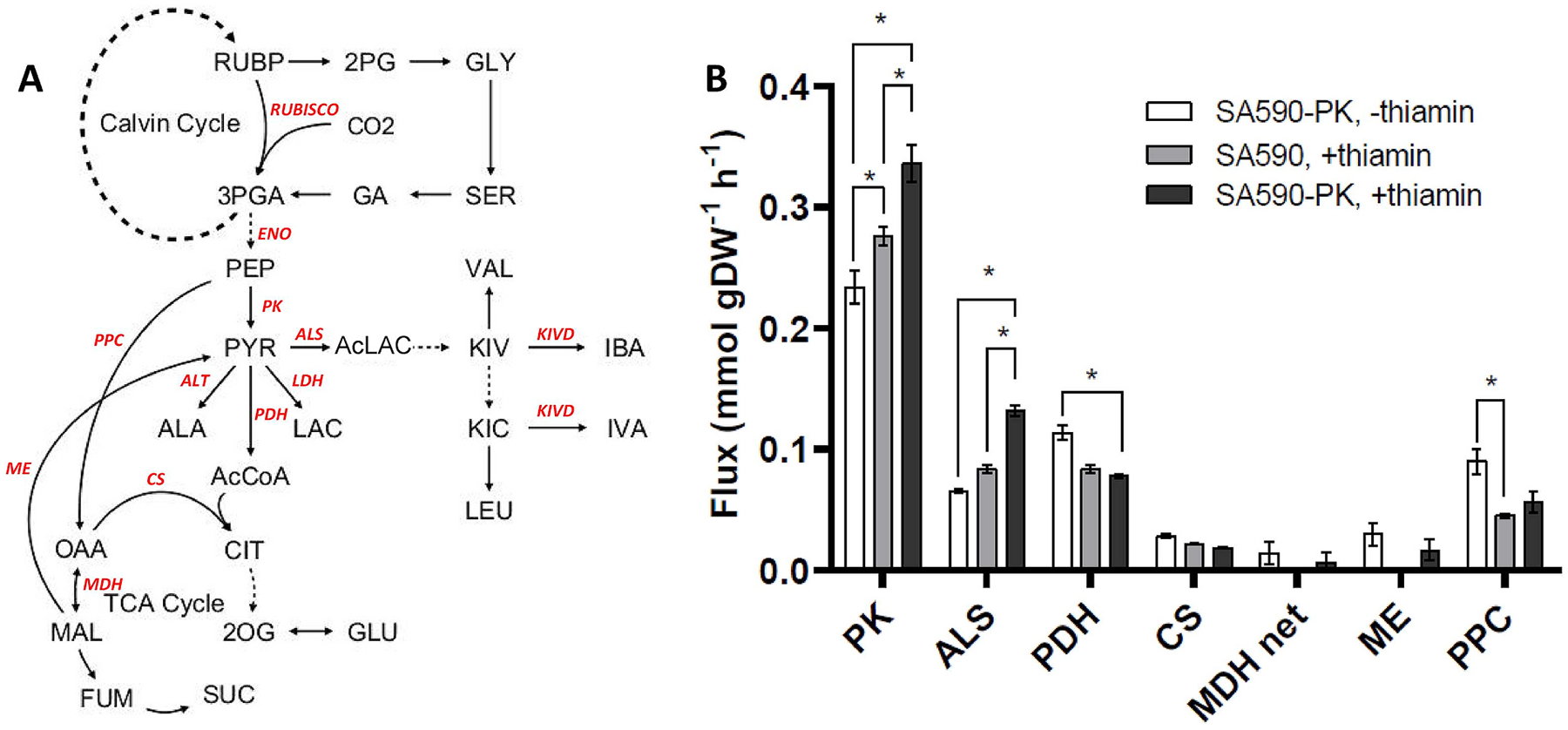
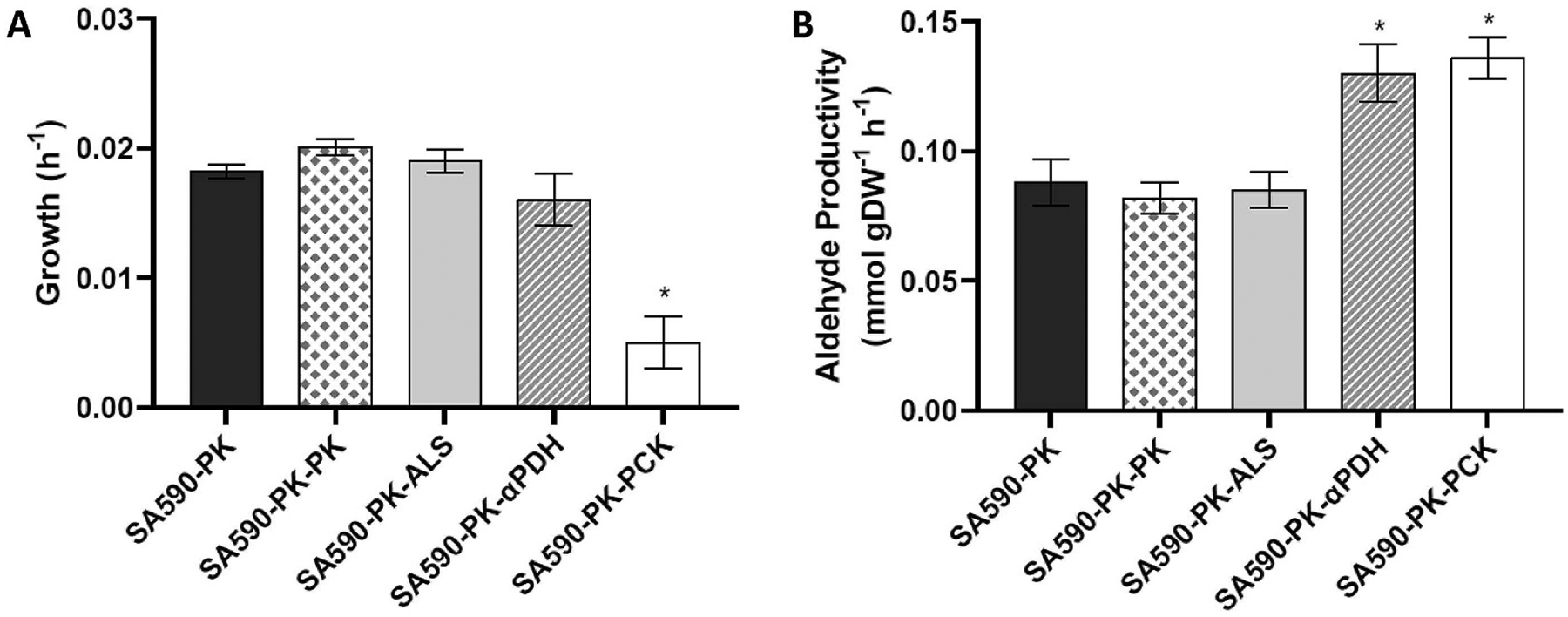
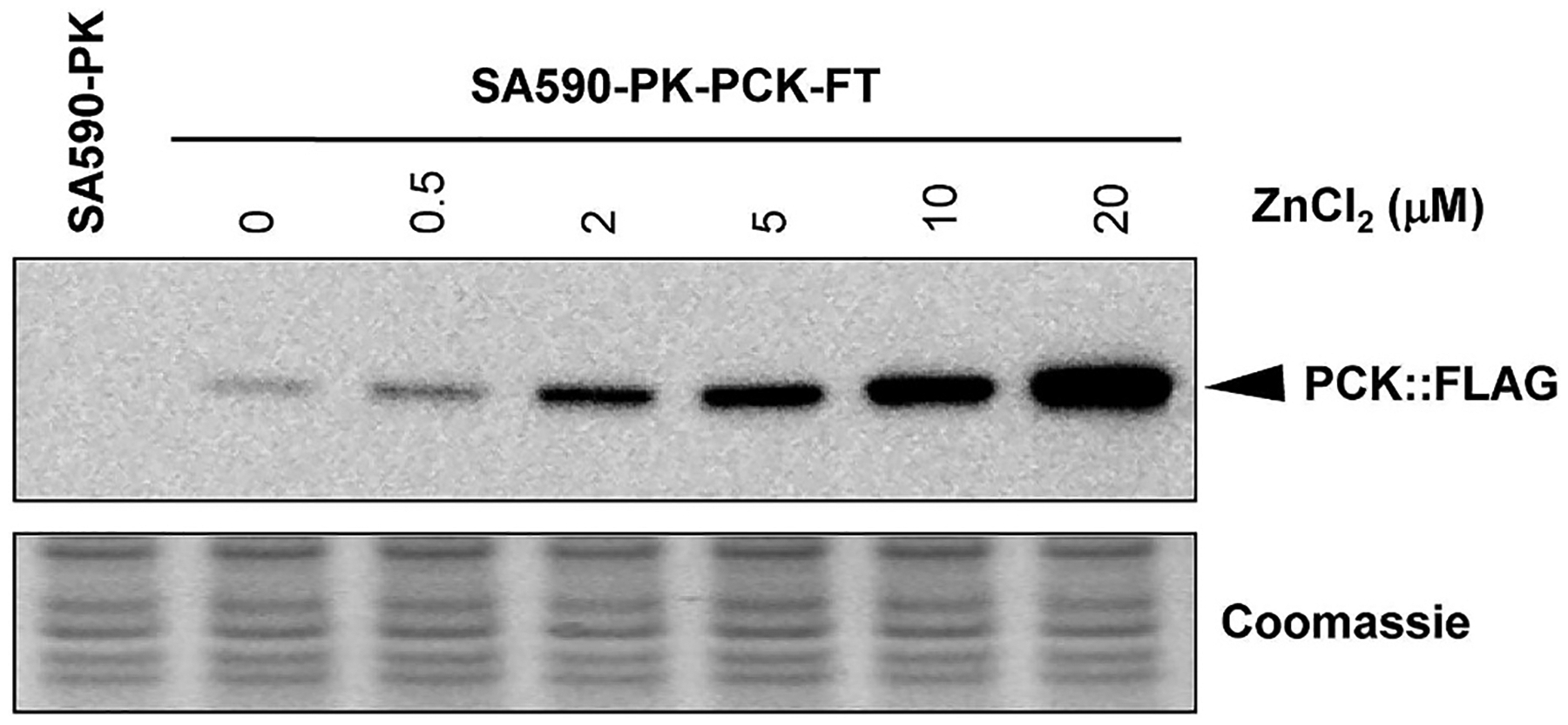
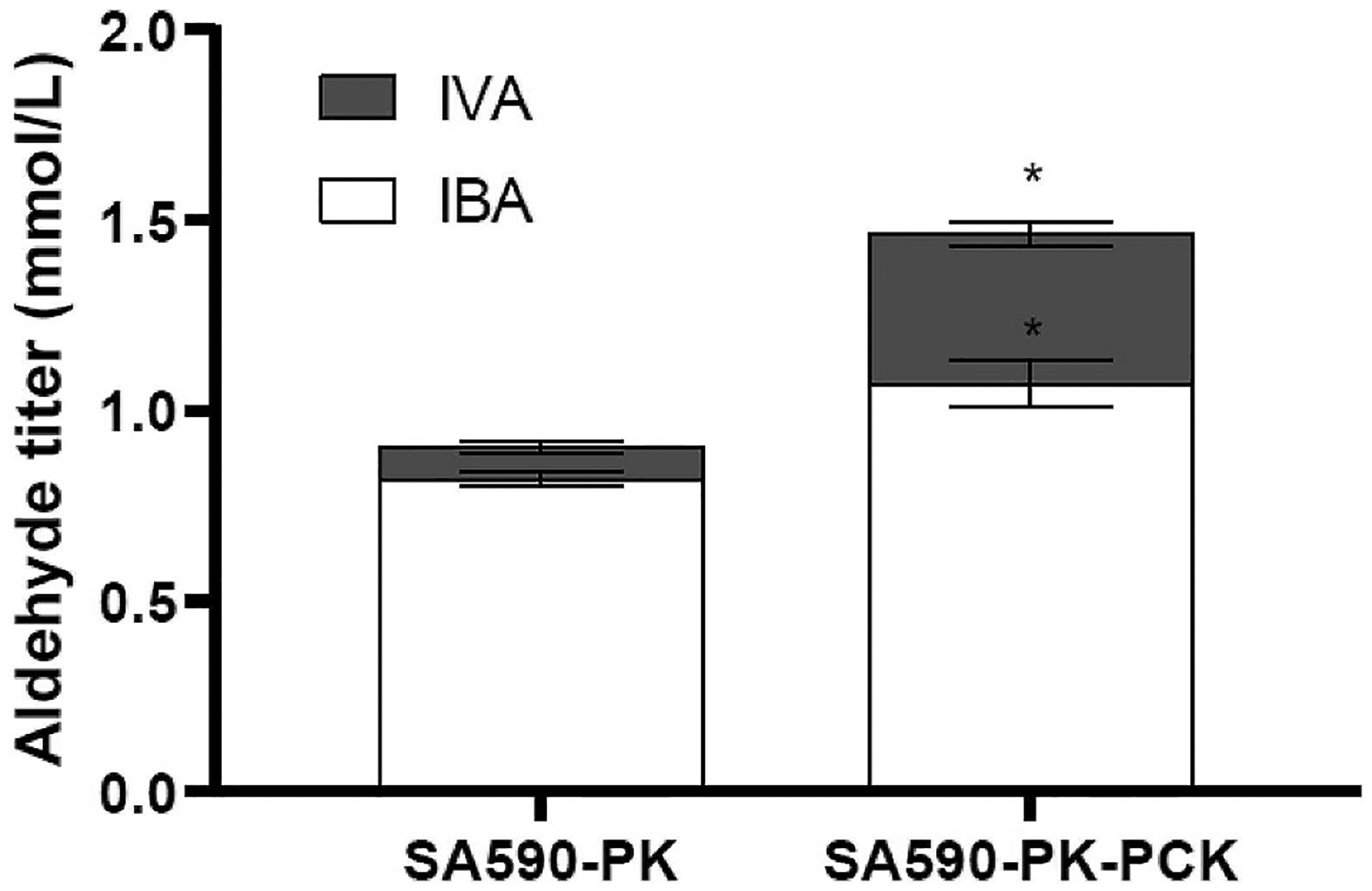
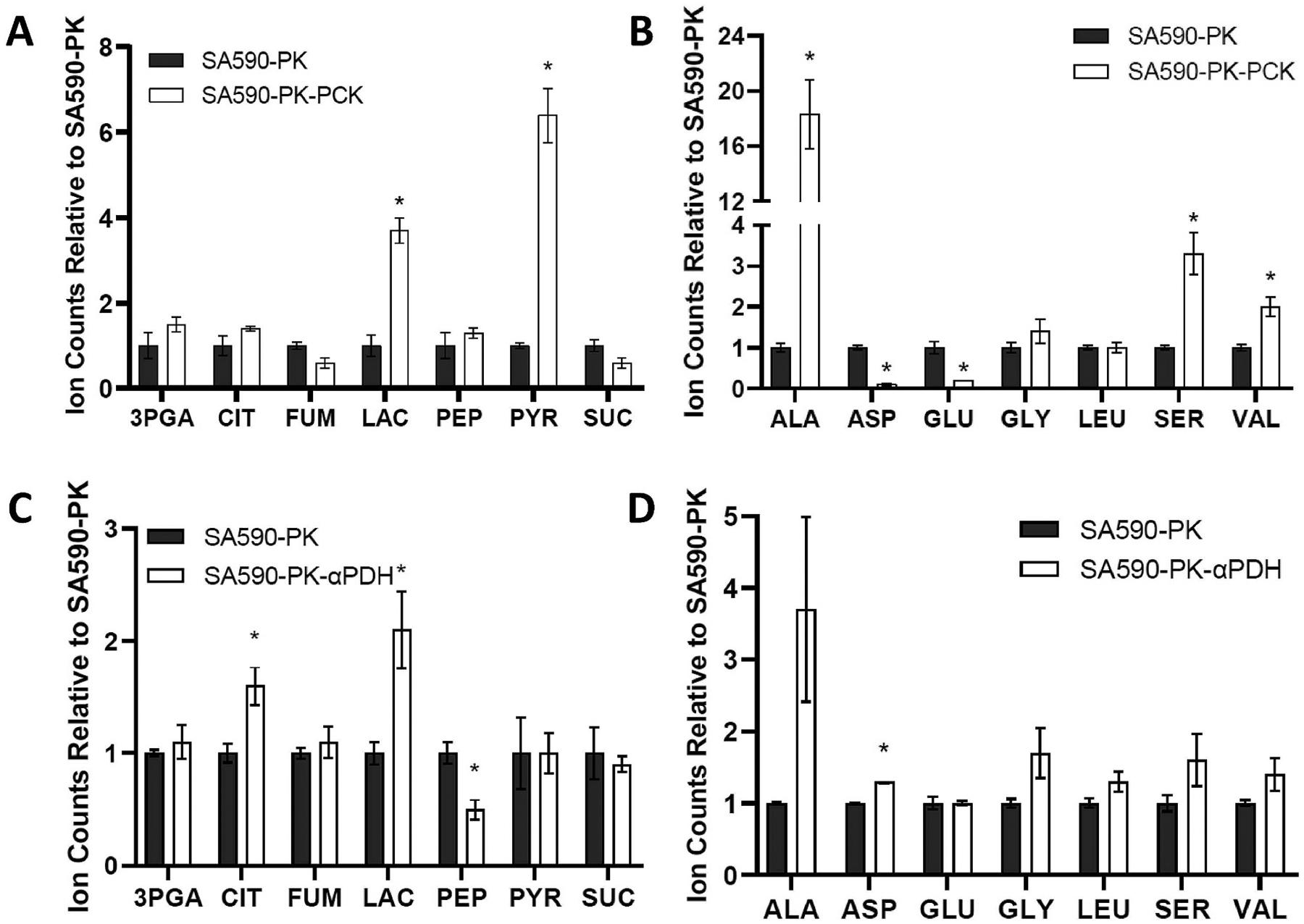
Isotopically nonstationary metabolic flux analysis (INST-MFA) provides a versatile platform to quantitatively assess in vivo metabolic activities of autotrophic systems. By applying INST-MFA to recombinant aldehyde-producing cyanobacteria, we identified metabolic alterations that correlated with increased strain performance in order to guide rational metabolic engineering. We identified four reactions adjacent to the pyruvate node that varied significantly with increasing aldehyde production: pyruvate kinase (PK) and acetolactate synthase (ALS) fluxes were directly correlated with product formation, while pyruvate dehydrogenase (PDH) and phosphoenolpyruvate carboxylase (PPC) fluxes were inversely correlated. Overexpression of enzymes for PK or ALS did not result in further improvements to the previous best-performing strain, while downregulation of PDH expression (through antisense RNA expression) or PPC flux (through expression of the reverse reaction, phosphoenolpyruvate carboxykinase) provided significant improvements. These results illustrate the potential of INST-MFA to enable a systematic approach for iterative identification and removal of pathway bottlenecks in autotrophic host cells.
Keywords: Cyanobacteria; Isobutyraldehyde; Metabolic flux analysis; Phosphoenolpyruvate; Photoautotrophic metabolism; Pyruvate; Stable isotope.
Copyright © 2020 International Metabolic Engineering Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Abernathy MH, Yu J, Ma F, Liberton M, Ungerer J, Hollinshead WD, Gopalakrishnan S, He L, Maranas CD, Pakrasi HB, Allen DK, Tang YJ, 2017. Deciphering cyanobacterial phenotypes for fast photoautotrophic growth via isotopically nonstationary metabolic flux analysis. Biotechnol. Biofuels 10, 273. - PMC - PubMed
-
- Adebiyi AO, Jazmin LJ, Young JD, 2015. 13C flux analysis of cyanobacterial metabolism. Photosynth. Res 126, 19–32. - PubMed
-
- Angermayr SA, Gorchs Rovira A, Hellingwerf KJ, 2015. Metabolic engineering of cyanobacteria for the synthesis of commodity products. Trends Biotechnol 33, 352–361. - PubMed
-
- Antoniewicz MR, 2013. 13C metabolic flux analysis: optimal design of isotopic labeling experiments. Curr. Opin. Biotechnol 24, 1116–1121. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
