

Bradykinin Receptors Play a Critical Role in the Chronic Post-ischaemia Pain Model
- PMID: 32222846
- PMCID: PMC11448614
- DOI: 10.1007/s10571-020-00832-3
Bradykinin Receptors Play a Critical Role in the Chronic Post-ischaemia Pain Model
Abstract
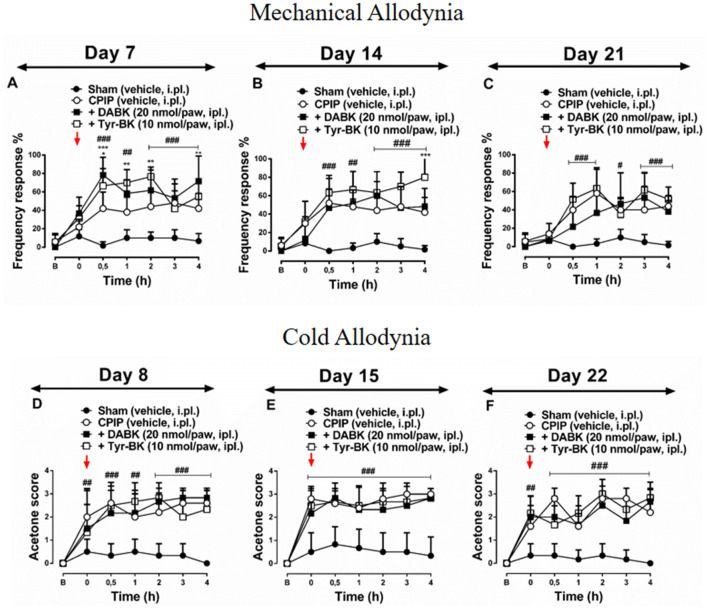
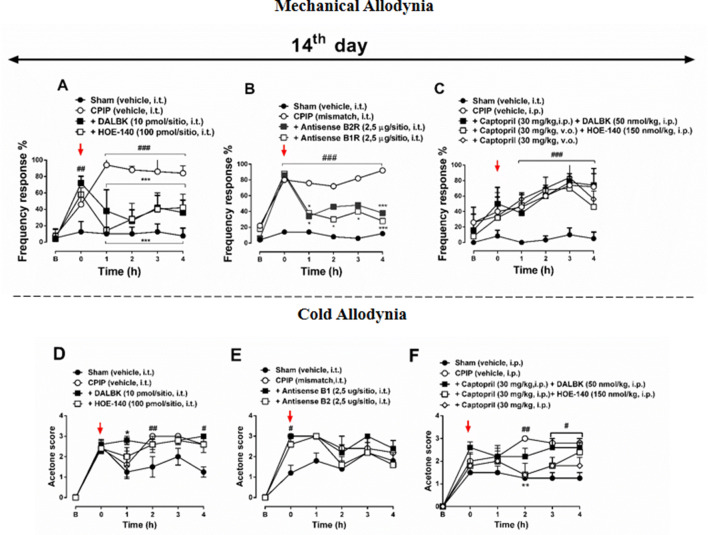
Complex regional pain syndrome type-I (CRPS-I) is a chronic painful condition resulting from trauma. Bradykinin (BK) is an important inflammatory mediator required in acute and chronic pain response. The objective of this study was to evaluate the association between BK receptors (B1 and B2) and chronic post-ischaemia pain (CPIP) development in mice, a widely accepted CRPS-I model. We assessed mechanical and cold allodynia, and paw oedema in male and female Swiss mice exposed to the CPIP model. Upon induction, the animals were treated with BKR antagonists (HOE-140 and DALBK); BKR agonists (Tyr-BK and DABK); antisense oligonucleotides targeting B1 and B2 and captopril by different routes in the model (7, 14 and 21 days post-induction). Here, we demonstrated that treatment with BKR antagonists, by intraperitoneal (i.p.), intraplantar (i.pl.), and intrathecal (i.t.) routes, mitigated CPIP-induced mechanical allodynia and oedematogenic response, but not cold allodynia. On the other hand, i.pl. administration of BKR agonists exacerbated pain response. Moreover, a single treatment with captopril significantly reversed the anti-allodynic effect of BKR antagonists. In turn, the inhibition of BKRs gene expression in the spinal cord inhibited the nociceptive behaviour in the 14th post-induction. The results of the present study suggest the participation of BKRs in the development and maintenance of chronic pain associated with the CPIP model, possibly linking them to CRPS-I pathogenesis.
Keywords: Bradykinin receptor antagonists; Chronic pain; Hyperalgesia; Inflammation; Inflammatory mediators; Kinins.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







References
-
- Beerthuizen A, Stronks DL, Van A et al (2012) Demographic and medical parameters in the development of complex regional pain syndrome type 1 (CRPS1): prospective study on 596 patients with a fracture. Pain 153:1187–1192 - PubMed
-
- Beirith A, Santos ARS, Calixto JB (2003) The role of neuropeptides and capsaicin-sensitive fibres in glutamate-induced nociception and paw oedema in mice. Brain Res 969:110–116 - PubMed
-
- Belichard P, Landry M, Faye P, Bachvarov DR (2000) Inflammatory hyperalgesia induced by zymosan in the plantar tissue of the rat : effect of kinin receptor antagonists. Immunopharmacology 46:139–147 - PubMed
-
- Borsook D, Sava S (2009) Pain: do ACE inhibitors exacerbate complex regional pain syndrome? Nat Rev Neurol 5:306–308 - PubMed
-
- Bortalanza B, Ferreira J, Hess SC (2002) Anti-allodynic action of the tormentic acid, a triterpene isolated from plant, against neuropathic and inflammatory persistent pain in mice. Eur J Pharmacol 453(453):203–208 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
