

Malaria exposure drives both cognate and bystander human B cells to adopt an atypical phenotype
- PMID: 32222961
- PMCID: PMC7611263
- DOI: 10.1002/eji.201948473
Malaria exposure drives both cognate and bystander human B cells to adopt an atypical phenotype
Abstract
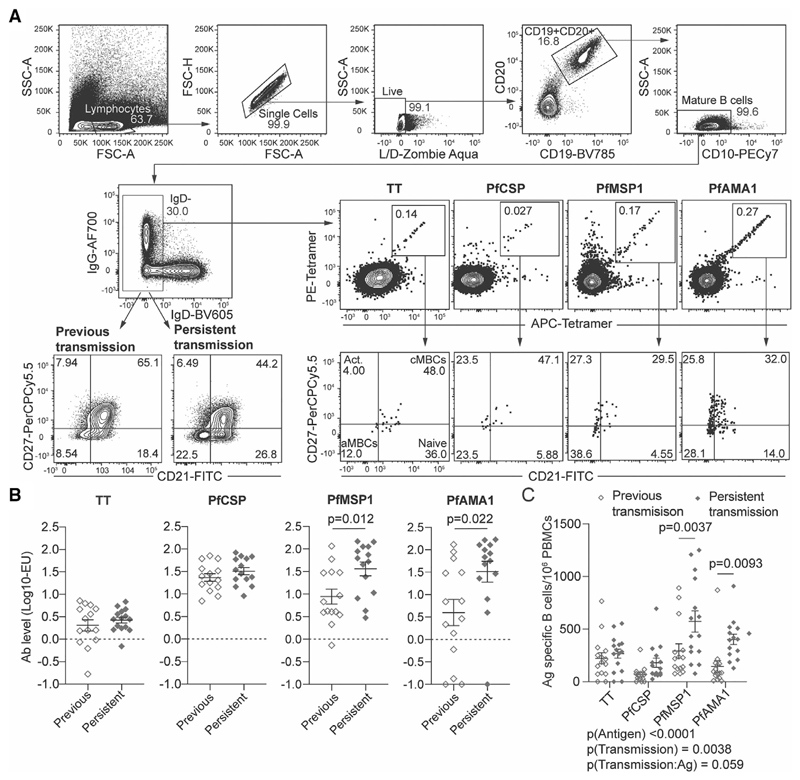
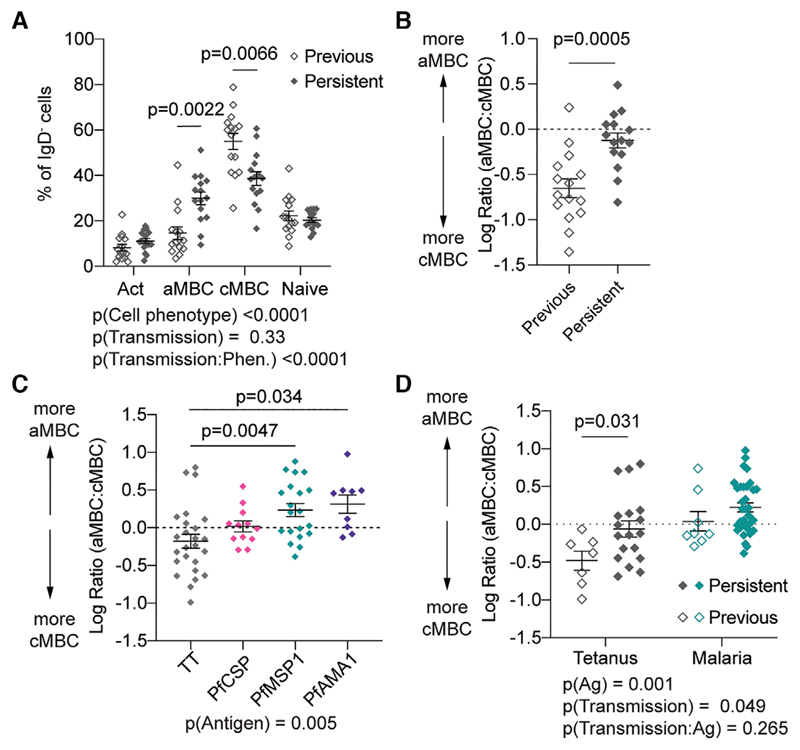
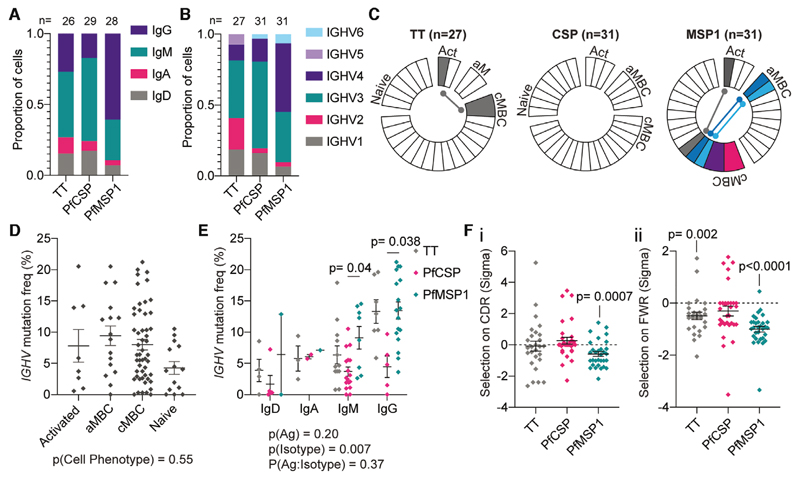
Atypical memory B cells (aMBCs) are found in elevated numbers in individuals exposed to malaria. A key question is whether malaria induces aMBCs as a result of exposure to Ag, or non-Ag-specific mechanisms. We identified Plasmodium and bystander tetanus toxoid (TT) specific B cells in individuals from areas of previous and persistent exposure to malaria using tetramers. Malaria-specific B cells were more likely to be aMBCs than TT-specific B cells. However, TT-specific B cells from individuals with continuous exposure to malaria were more likely to be aMBCs than TT-specific B cells in individuals from areas where transmission has ceased. Finally, sequences of BCRs specific for a blood stage malaria-Ag were more highly mutated than sequences from TT-specific BCRs and under strong negative selection, indicative of ongoing antigenic pressure. Our data suggest both persistent Ag exposure and the inflammatory environment shape the B-cell response to malaria and bystander Ags.
Keywords: B-cell memory; Plasmodium; immunological memory; malaria; tetanus toxoid.
© 2020 The Authors. European Journal of Immunology published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
Figures
References
-
- World Health Organization, World Malaria Report 2019. World Health Organization. Geneva: 2019.
-
- Langhorne J, Ndungu FM, Sponaas AM, Marsh K. Immunity to malaria: more questions than answers. Nat Immunol. 2008;9:725–732. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
