

A branched heterochronic pathway directs juvenile-to-adult transition through two LIN-29 isoforms
- PMID: 32223899
- PMCID: PMC7105380
- DOI: 10.7554/eLife.53387
A branched heterochronic pathway directs juvenile-to-adult transition through two LIN-29 isoforms
Abstract
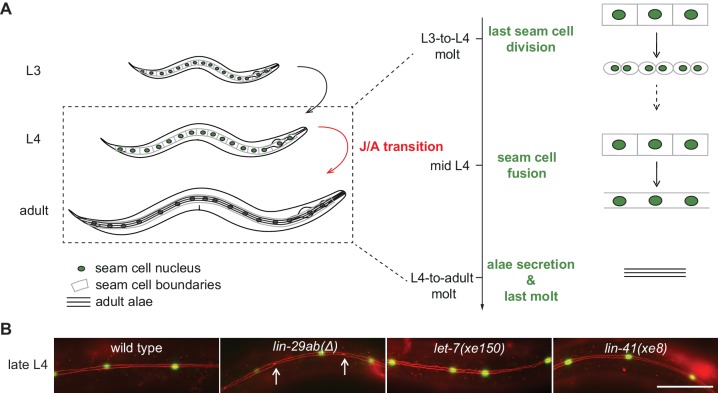
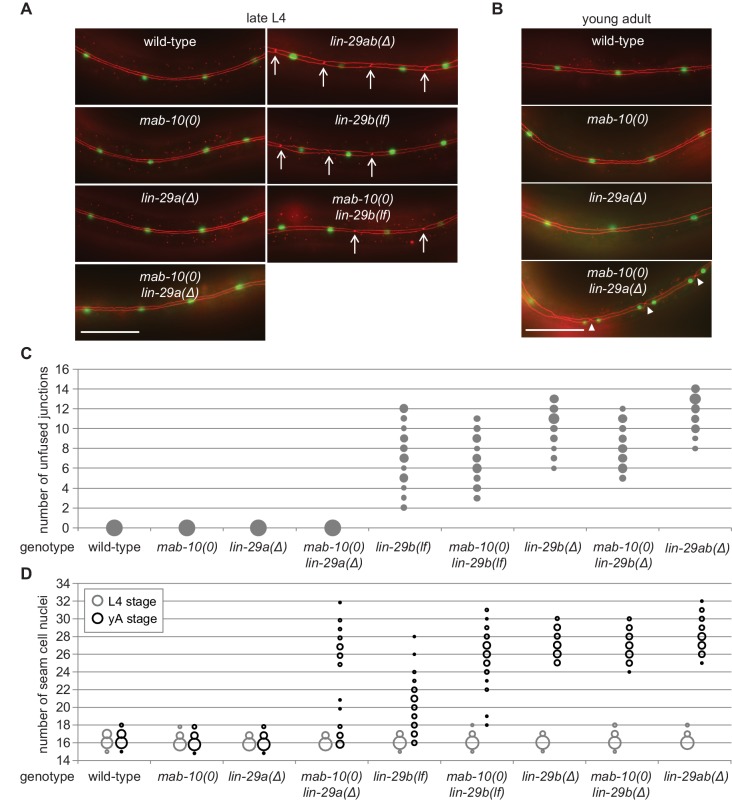
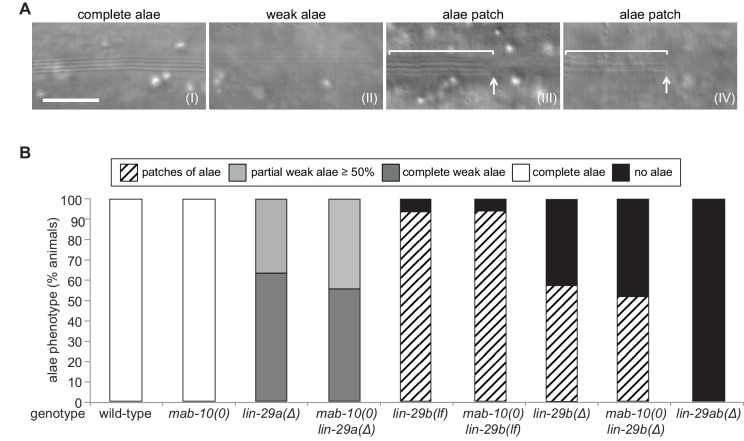
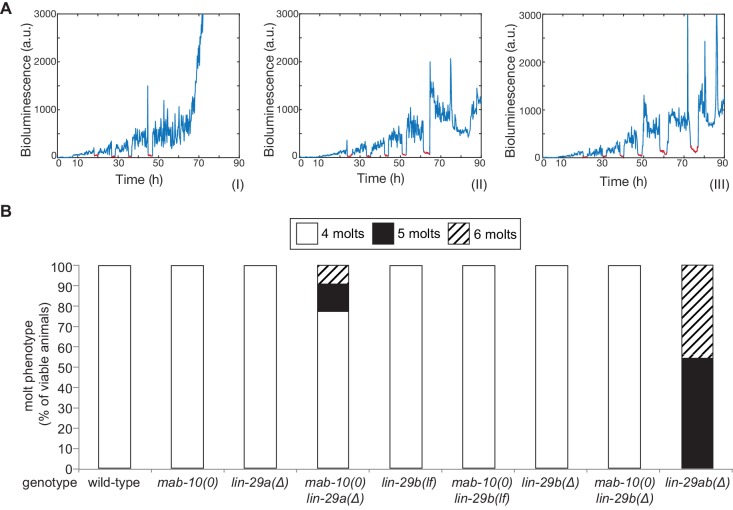
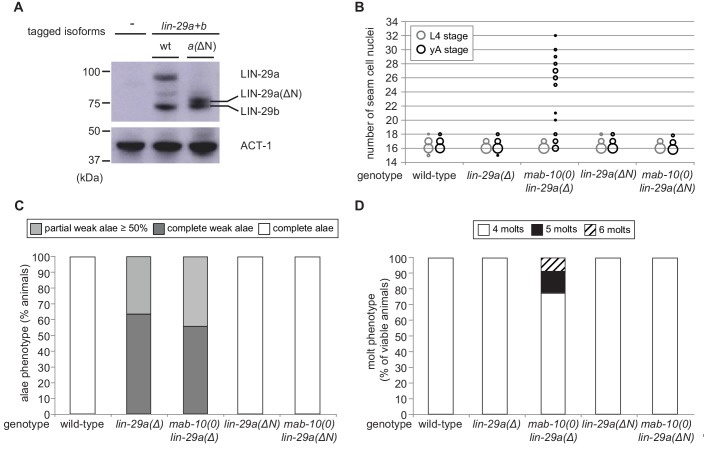
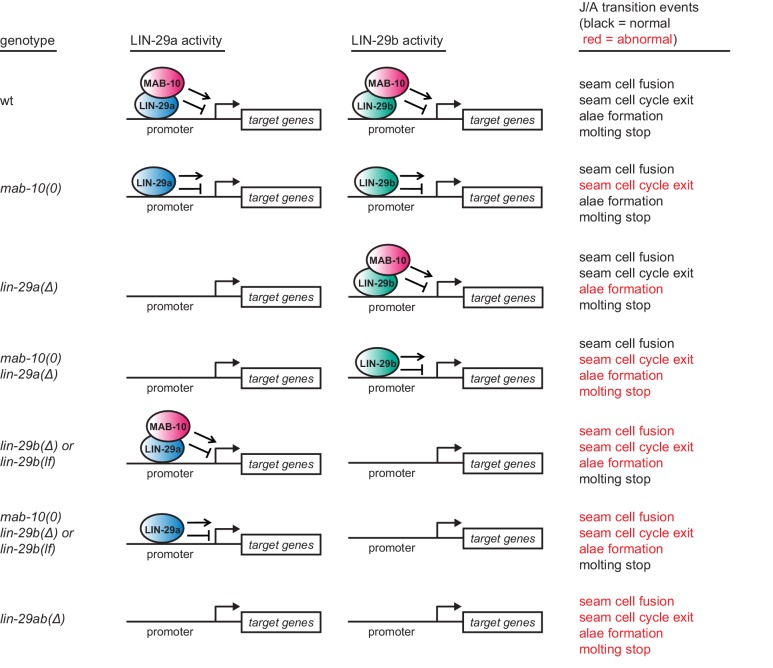
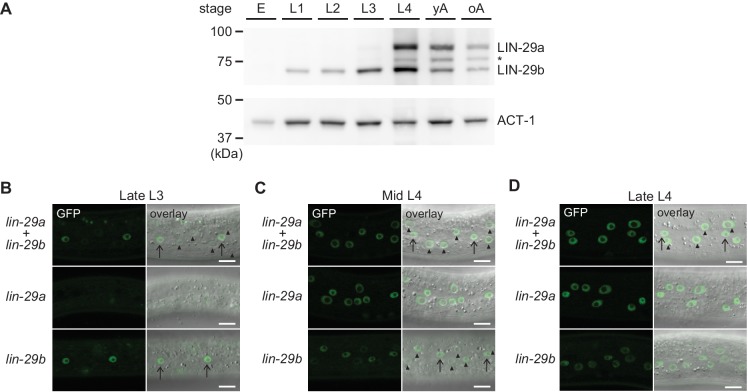
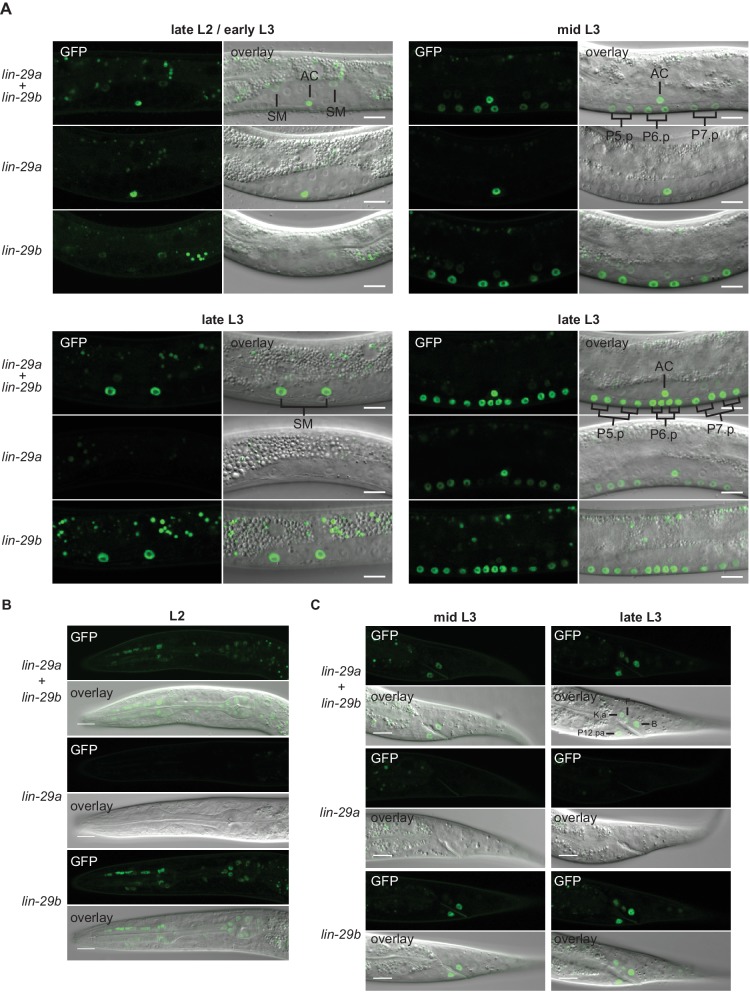
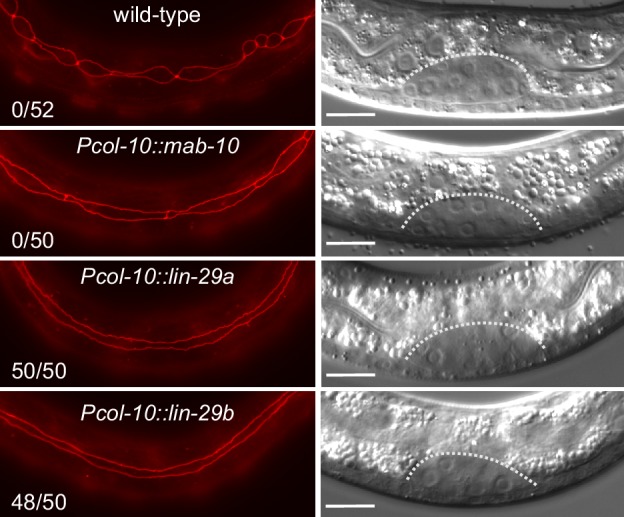
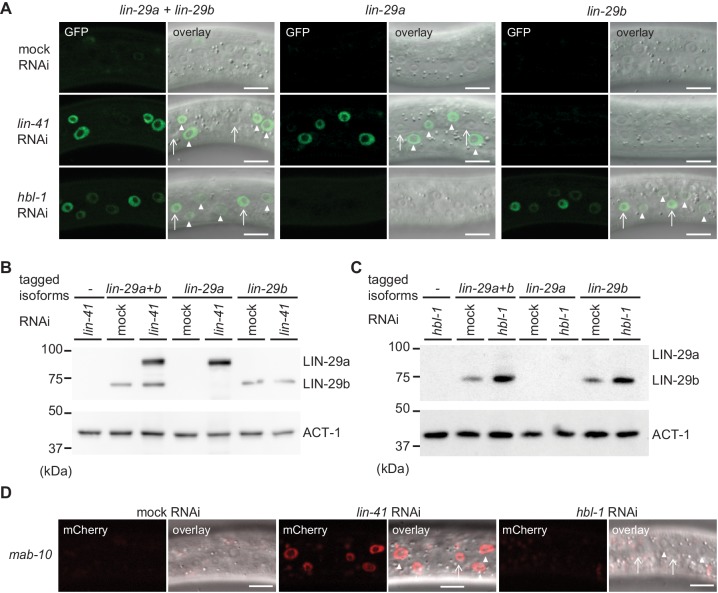
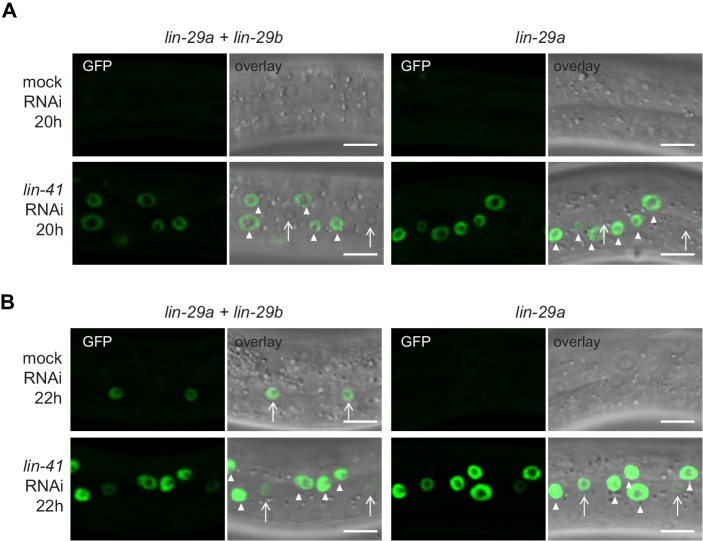
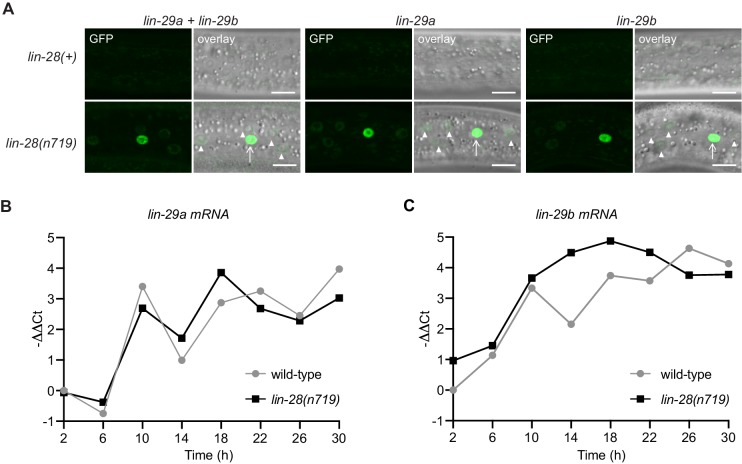
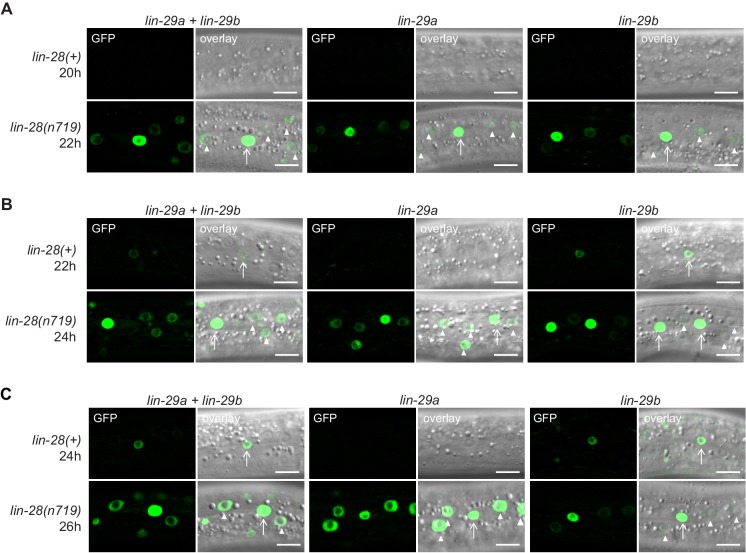
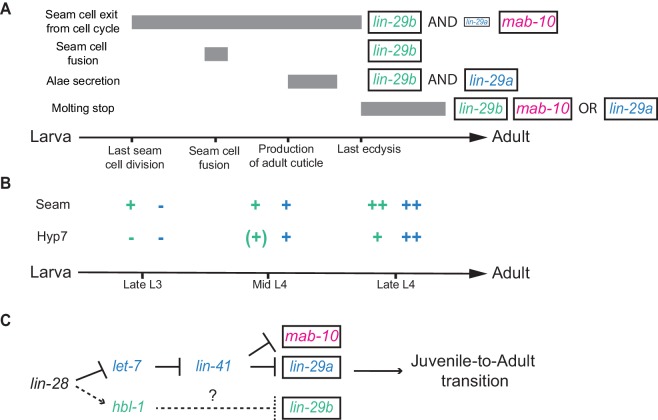
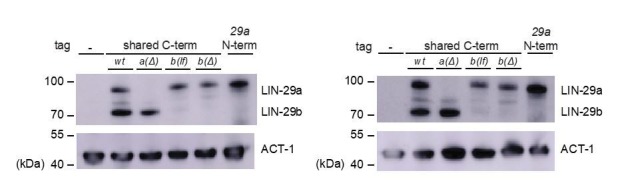
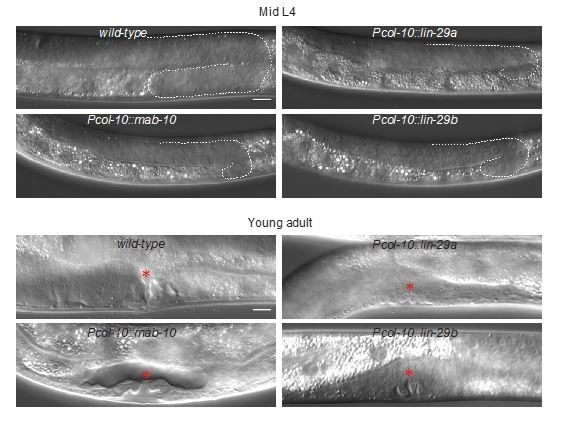
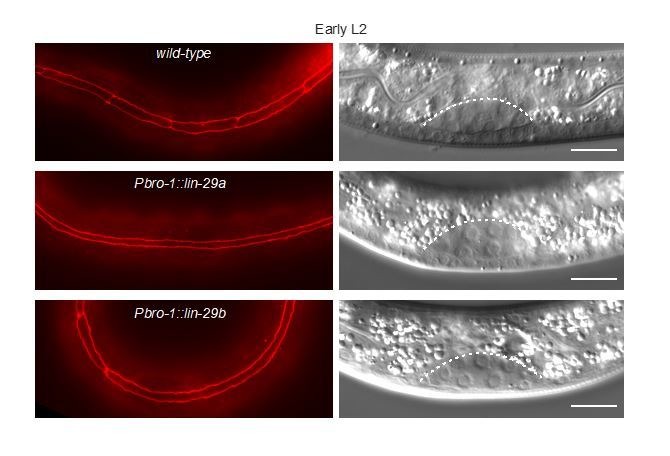
Robust organismal development relies on temporal coordination of disparate physiological processes. In Caenorhabditis elegans, the heterochronic pathway controls a timely juvenile-to-adult (J/A) transition. This regulatory cascade of conserved proteins and small RNAs culminates in accumulation of the transcription factor LIN-29, which triggers coordinated execution of transition events. We report that two LIN-29 isoforms fulfill distinct functions. Functional specialization is a consequence of distinct isoform expression patterns, not protein sequence, and we propose that distinct LIN-29 dose sensitivities of the individual J/A transition events help to ensure their temporal ordering. We demonstrate that unique isoform expression patterns are generated by the activities of LIN-41 for lin-29a, and of HBL-1 for lin-29b, whereas the RNA-binding protein LIN-28 coordinates LIN-29 isoform activity, in part by regulating both hbl-1 and lin-41. Our findings reveal that coordinated transition from juvenile to adult involves branching of a linear pathway to achieve timely control of multiple events.
Keywords: C. elegans; developmental biology; developmental timing; epidermis; heterochronic; molt; puberty; terminal differentiation.
© 2020, Azzi et al.
Conflict of interest statement
CA, AN, HG No competing interests declared, FA The author is now affiliated with CSL Behring, Research, CSL Biologics Research Center, however all work was conducted when affiliated with Friedrich Miescher Institute for Biomedical Research and University of Basel.
Figures



References
-
- Abbott AL, Alvarez-Saavedra E, Miska EA, Lau NC, Bartel DP, Horvitz HR, Ambros V. The let-7 MicroRNA family members mir-48, mir-84, and mir-241 function together to regulate developmental timing in Caenorhabditis elegans. Developmental Cell. 2005;9:403–414. doi: 10.1016/j.devcel.2005.07.009. - DOI - PMC - PubMed
-
- Abete-Luzi P, Fukushige T, Yun S, Krause MW, Eisenmann DM. New roles for the heterochronic transcription factor LIN-29 in cuticle maintenance and lipid metabolism at the Larval-to-Adult transition in Caenorhabditis elegans. Genetics. 2020;214:669–690. doi: 10.1534/genetics.119.302860. - DOI - PMC - PubMed
-
- Abreu AP, Dauber A, Macedo DB, Noel SD, Brito VN, Gill JC, Cukier P, Thompson IR, Navarro VM, Gagliardi PC, Rodrigues T, Kochi C, Longui CA, Beckers D, de Zegher F, Montenegro LR, Mendonca BB, Carroll RS, Hirschhorn JN, Latronico AC, Kaiser UB. Central precocious puberty caused by mutations in the imprinted gene MKRN3. New England Journal of Medicine. 2013;368:2467–2475. doi: 10.1056/NEJMoa1302160. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
