

Wild-type IDH2 contributes to Epstein-Barr virus-dependent metabolic alterations and tumorigenesis
- PMID: 32224436
- PMCID: PMC7109632
- DOI: 10.1016/j.molmet.2020.02.009
Wild-type IDH2 contributes to Epstein-Barr virus-dependent metabolic alterations and tumorigenesis
Abstract
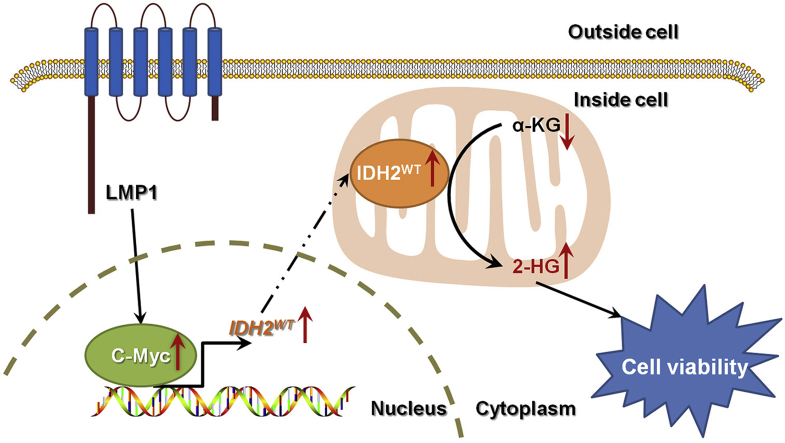
Objective: Epstein-Barr virus (EBV) is a well-recognized oncogenic virus that can induce host cell metabolic reprogramming and tumorigenesis by targeting vital metabolic enzymes or regulators. This study aims to explore the role of wild-type isocitrate dehydrogenase 2 (IDH2) in metabolic reprogramming and tumorigenesis induced by EBV-encoded latent membrane protein 1 (LMP1).
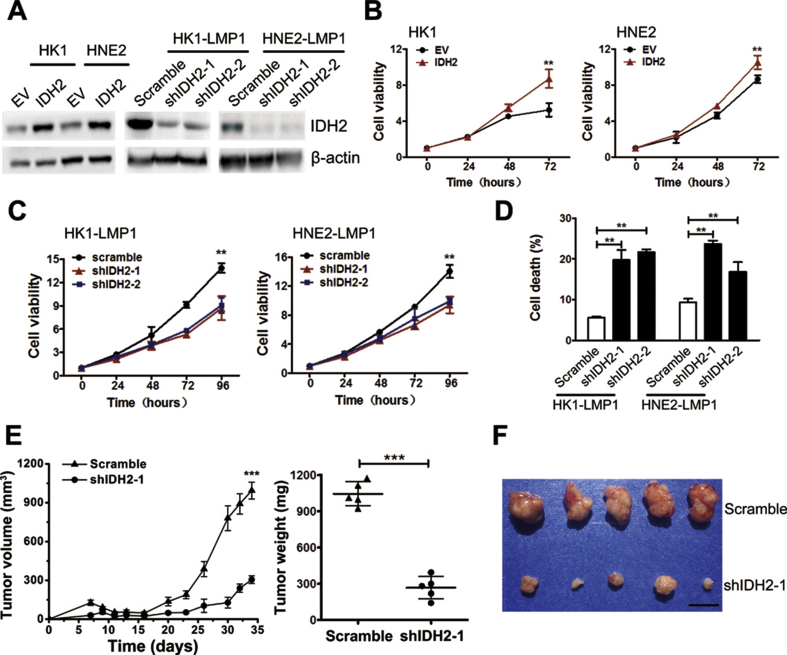
Methods: Mechanistic dissection of wild-type IDH2 in EBV-LMP1-induced tumorigenesis was investigated using western blotting, real-time polymerase chain reaction (PCR), immunochemistry, chromatin immunoprecipitation (ChIP), and luciferase assay. The role of wild-type IDH2 was examined by cell viability assays/Sytox Green staining in vitro and xenograft assays in vivo.
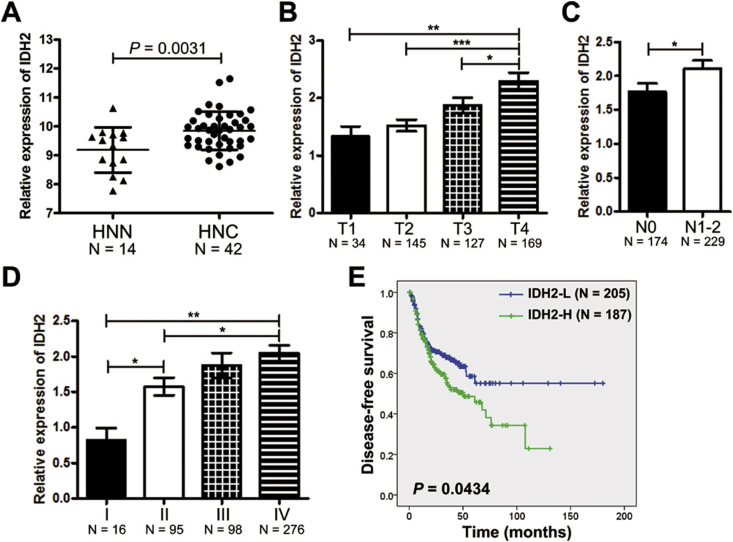
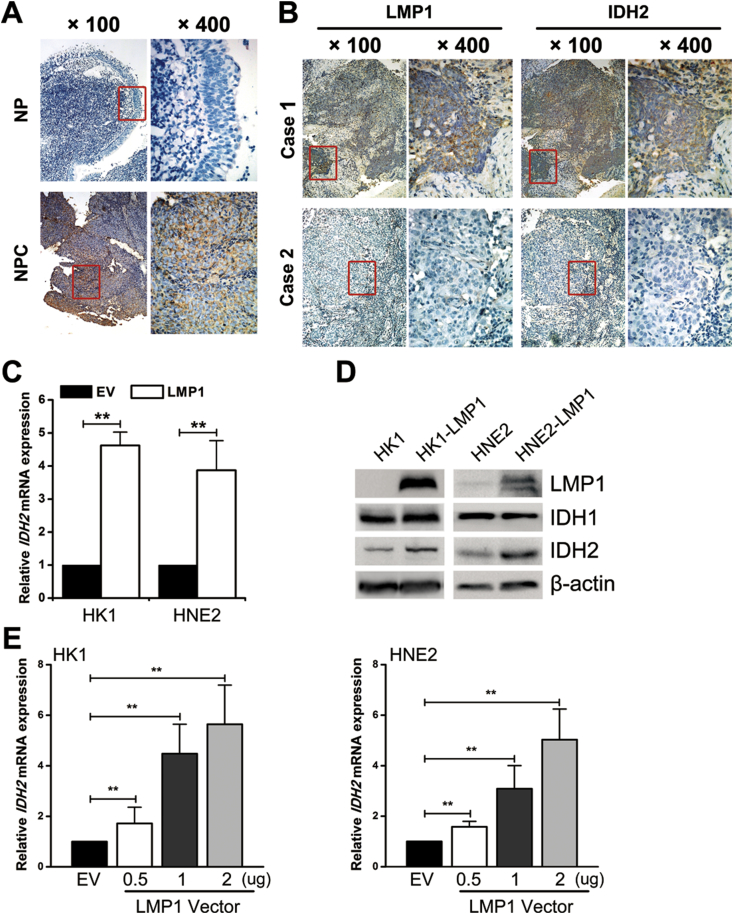
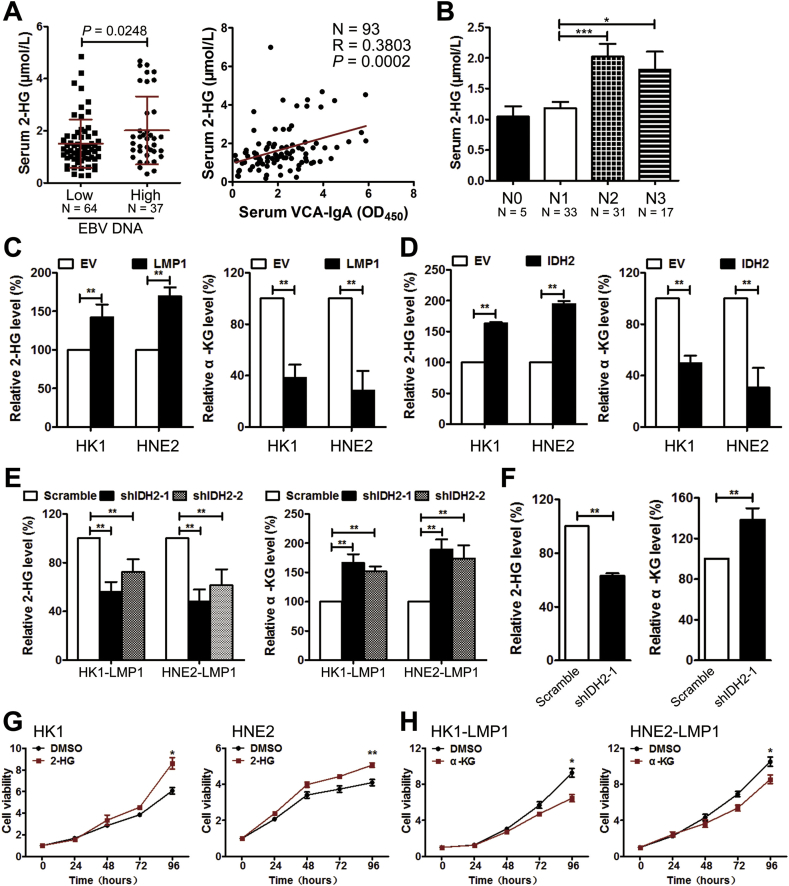
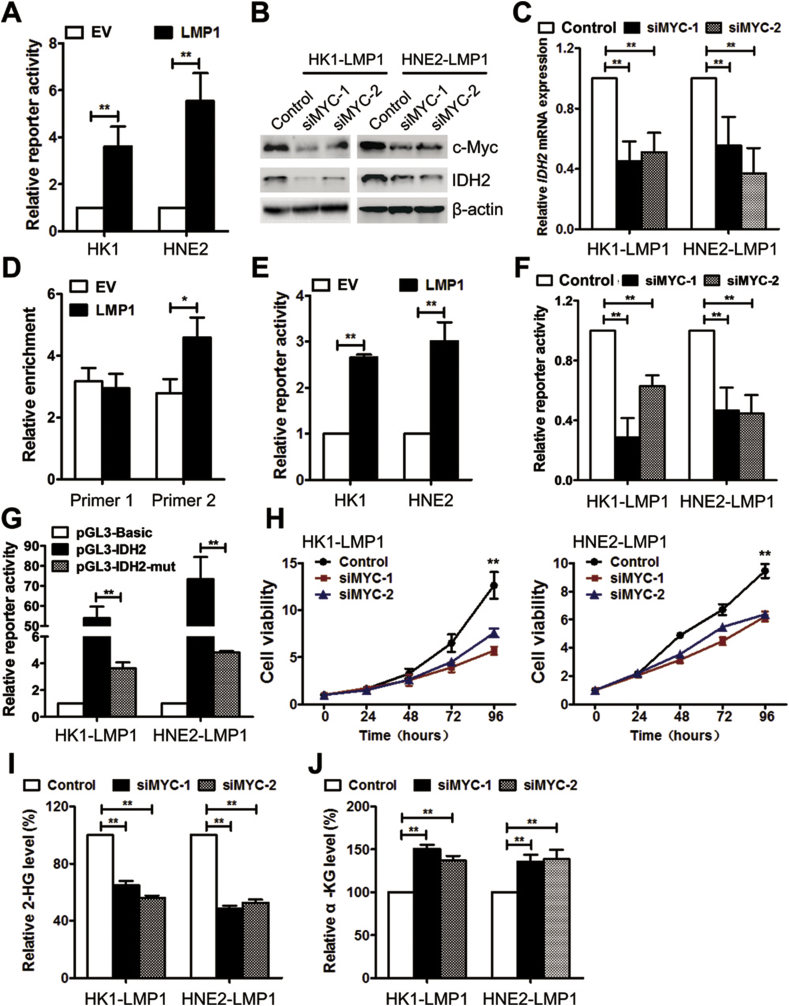
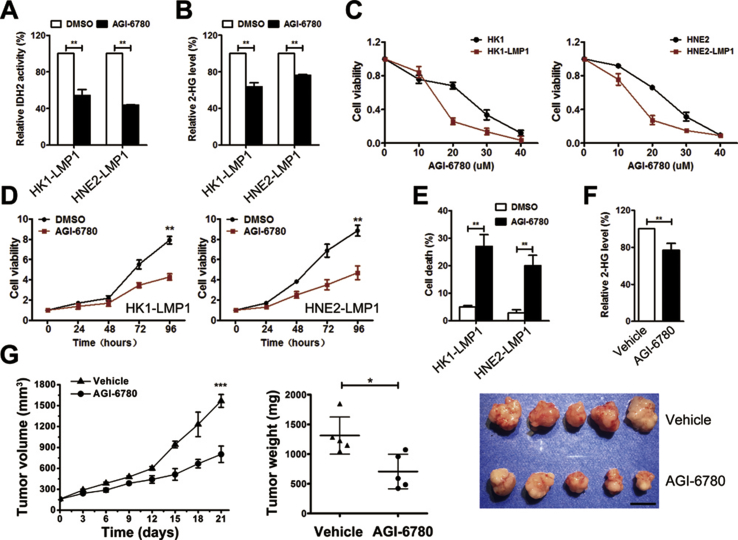
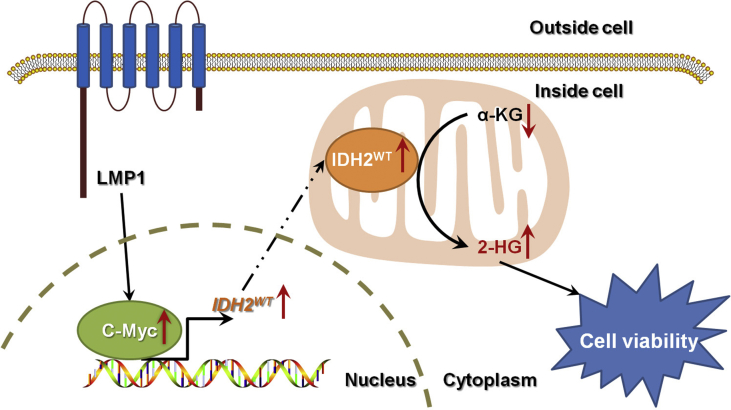
Results: IDH2 over-expression is a prognostic indicator of poorer disease-free survival for patients with head and neck squamous cell carcinoma (HNSCC). IDH2 expression is also upregulated in nasopharyngeal carcinoma (NPC, a subtype of HNSCC) tissues, which is positively correlated with EBV-LMP1 expression. EBV-LMP1 contributes to NPC cell viability and xenograft tumor growth mediated through wild-type IDH2. IDH2-dependent changes in intracellular α-ketoglutarate (α-KG) and 2-hydroxyglutarate (2-HG) contribute to EBV-LMP1-induced tumorigenesis in vitro and in vivo. Elevated serum 2-HG level is associated with high EBV DNA and viral capsid antigen-immunoglobulin A (VCA-IgA) levels in patients with NPC. A significantly positive correlation exists between serum 2-HG level and regional lymph node metastases of NPC. EBV-LMP1 enhances the binding of c-Myc with the IDH2 promoter and transcriptionally activates wild-type IDH2 through c-Myc. Targeting IDH2 decreased intracellular 2-HG levels and survival of EBV-LMP1-positive tumor cells in vitro and in vivo.
Conclusions: Our results demonstrate that the EBV-LMP1/c-Myc/IDH2WT signaling axis is critical for EBV-dependent metabolic changes and tumorigenesis, which may provide new insights into EBV-related cancer diagnosis and therapy.
Keywords: Epstein-Barr virus; Isocitrate dehydrogenase 2; Metabolic reprogramming; Nasopharyngeal carcinoma; Tumorigenesis.
Copyright © 2020. Published by Elsevier GmbH.
Figures
References
-
- Plummer M., de Martel C., Vignat J., Ferlay J., Bray F., Franceschi S. Global burden of cancers attributable to infections in 2012: a synthetic analysis. Lancet Global Health. 2016;4(9):e609–e616. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
