

The Role of Hydrogen Peroxide and Peroxiredoxins throughout the Cell Cycle
- PMID: 32224940
- PMCID: PMC7222192
- DOI: 10.3390/antiox9040280
The Role of Hydrogen Peroxide and Peroxiredoxins throughout the Cell Cycle
Abstract
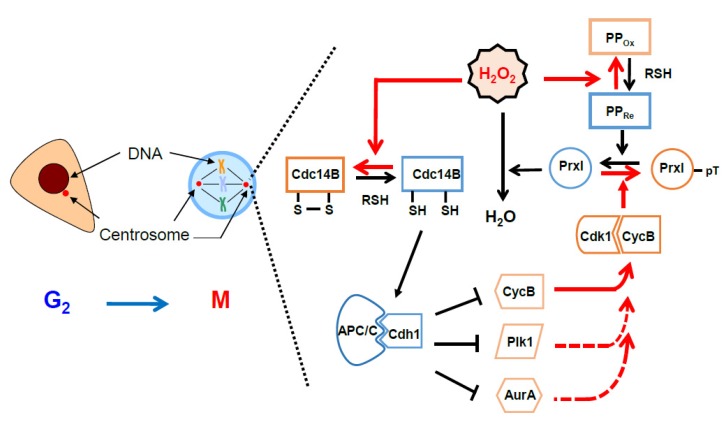
Hydrogen peroxide (H2O2) is an oxidizing agent that induces cellular damage at inappropriate concentrations and gives rise to an arrest during cell cycle progression, causing cell death. Recent evidence indicates that H2O2 also acts as a promoter for cell cycle progression by oxidizing specific thiol proteins. The intracellular concentration of H2O2 is regulated tightly, enabling its use as a cellular signaling molecule while minimizing its potential to cause cellular damage. Peroxiredoxins (Prxs) have peroxidase activity toward H2O2, organic hydroperoxides, and peroxynitrite for protecting cells from oxidative stress. They are suggested to work as signaling mediators, allowing the local accumulation of H2O2 by inactivating their peroxidase activity uniquely compared with other antioxidant proteins such as catalase and glutathione peroxidase. Given that Prxs are highly sensitive to oxidation by H2O2, they act as sensors and transducers of H2O2 signaling via transferring their oxidation state to effector proteins. The concentrations of intracellular H2O2 increase as the cell cycle progresses from G1 to mitosis. Here, we summarize the roles of Prxs with regard to the regulation of cell cycle-dependent kinase activity and anaphase-promoting complex/cyclosome in terms of changes in H2O2 levels. Protection of the cell from unwanted progression of the cell cycle is suggested to be a role of Prx. We discuss the possible roles of Prxs to control H2O2 levels.
Keywords: cell cycle; centrosome; cyclin-dependent kinase; hydrogen peroxide; peroxiredoxin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
Grants and funding
- 2019R1A6C1010020/National Research Foundation of Korea
- 2018R1D1A1A02049371/National Research Foundation of Korea
- 2019R1A5A6099645/National Research Foundation of Korea
- NRF-2015-Fostering Core Leaders of the Future Basic Science Program/Global Ph.D. Fellowship Program/National Research Foundation of Korea
LinkOut - more resources
Full Text Sources
