

The isoleucic acid triad: distinct impacts on plant defense, root growth, and formation of reactive oxygen species
- PMID: 32227083
- PMCID: PMC7448199
- DOI: 10.1093/jxb/eraa160
The isoleucic acid triad: distinct impacts on plant defense, root growth, and formation of reactive oxygen species
Abstract
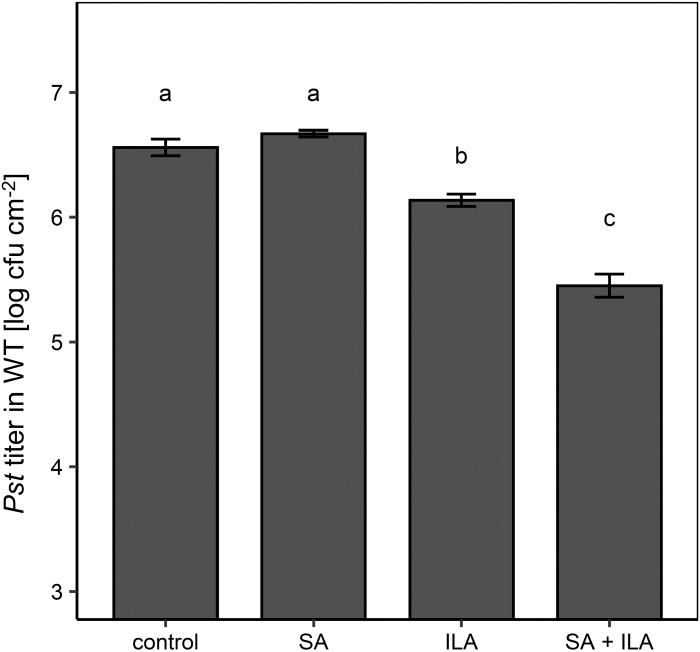
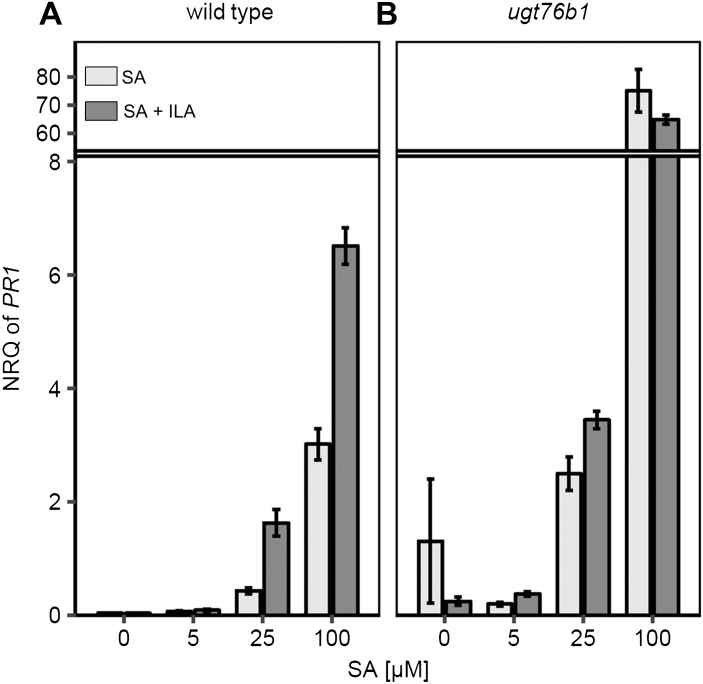
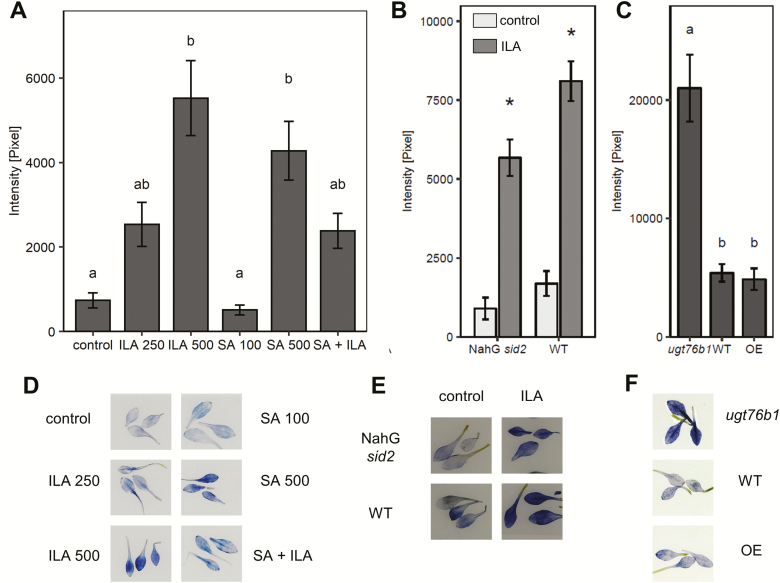
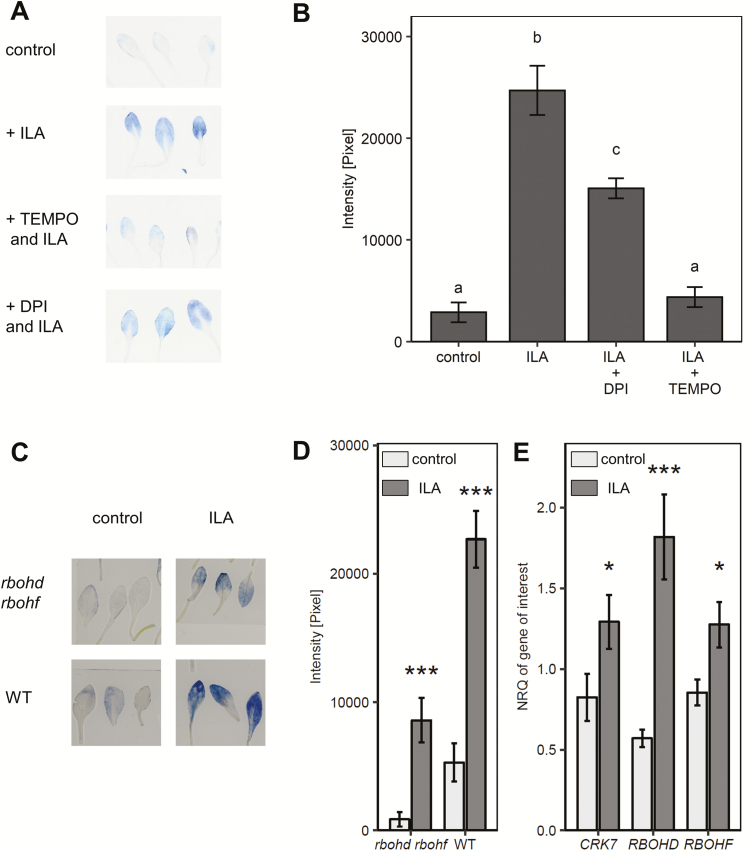
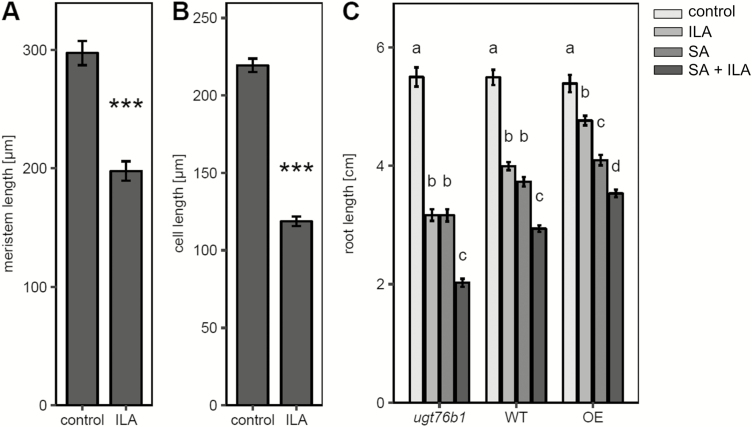
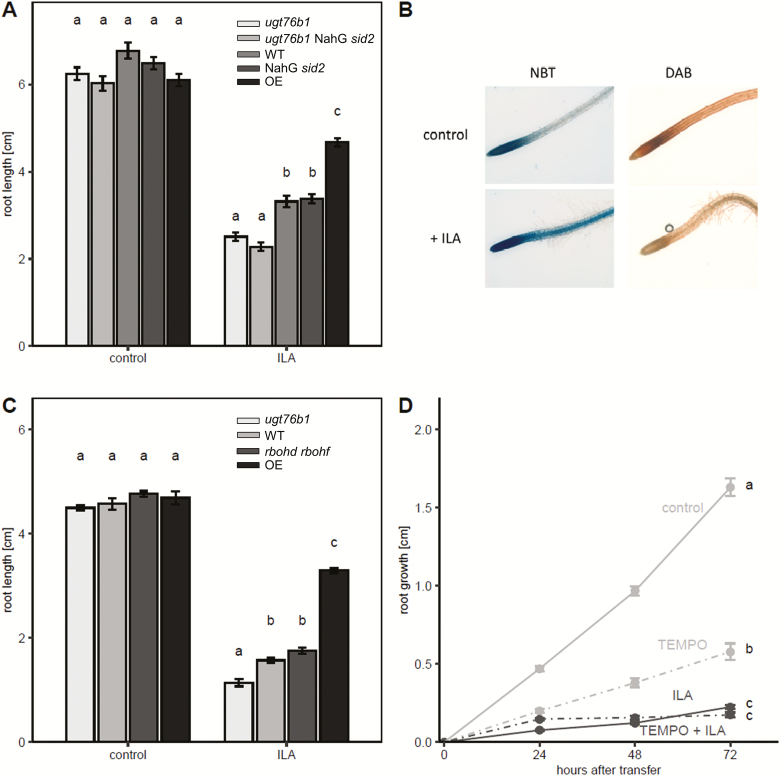
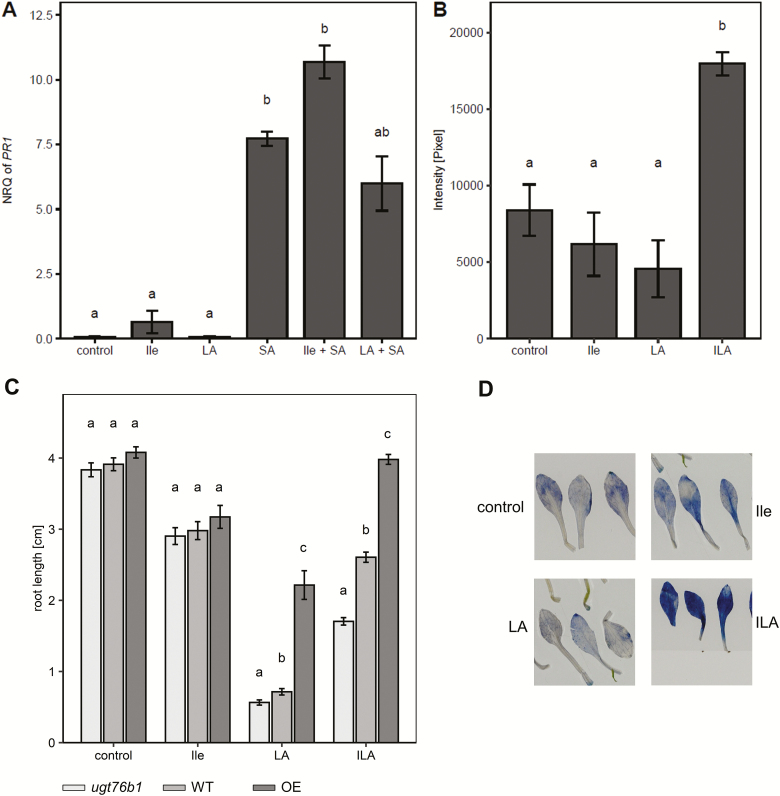
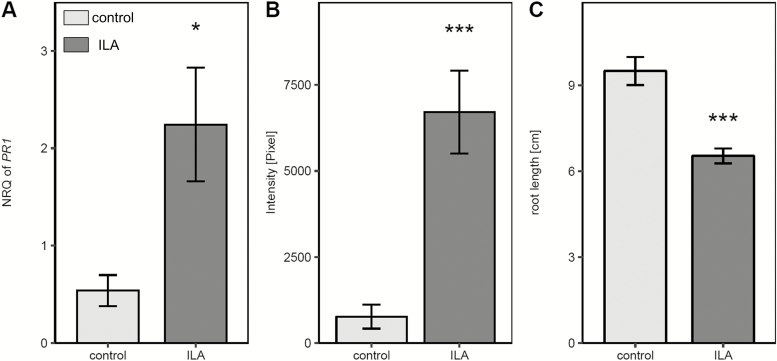
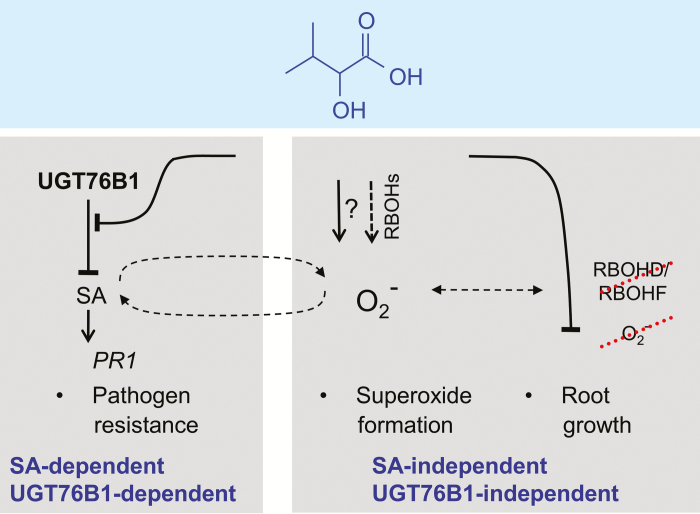
Isoleucic acid (ILA), a branched-chain amino acid-related 2-hydroxycarboxylic acid, occurs ubiquitously in plants. It enhances pathogen resistance and inhibits root growth of Arabidopsis. The salicylic acid (SA) glucosyltransferase UGT76B1 is able to conjugate ILA. Here, we investigate the role of ILA in planta in Arabidopsis and reveal a triad of distinct responses to this small molecule. ILA synergistically co-operates with SA to activate SA-responsive gene expression and resistance in a UGT76B1-dependent manner in agreement with the observed competitive ILA-dependent repression of SA glucosylation by UGT76B1. However, ILA also shows an SA-independent stress response. Nitroblue tetrazolium staining and pharmacological experiments indicate that ILA induces superoxide formation of the wild type and of an SA-deficient (NahG sid2) line. In contrast, the inhibitory effect of ILA on root growth is independent of both SA and superoxide induction. These effects of ILA are specific and distinct from its isomeric compound leucic acid and from the amino acid isoleucine. Leucic acid and isoleucine do not induce expression of defense marker genes or superoxide production, whereas both compounds inhibit root growth. All three responses to ILA are also observed in Brassica napus.
Keywords: Brassica napus; Arabidopsis; glucosyltransferase; isoleucic acid; plant defense; reactive oxygen species; root growth; salicylic acid; superoxide anion.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Alonso JM, Stepanova AN, Leisse TJ, et al. 2003. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653–657. - PubMed
-
- Chen J, Zhang Y, Wang C, Lü W, Jin JB, Hua X. 2011. Proline induces calcium-mediated oxidative burst and salicylic acid signaling. Amino Acids 40, 1473–1484. - PubMed
-
- Chen X, Truksa M, Shah S, Weselake RJ. 2010. A survey of quantitative real-time polymerase chain reaction internal reference genes for expression studies in Brassica napus. Analytical Biochemistry 405, 138–140. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
