

The transition to compulsion in addiction
- PMID: 32231315
- PMCID: PMC7610550
- DOI: 10.1038/s41583-020-0289-z
The transition to compulsion in addiction
Abstract
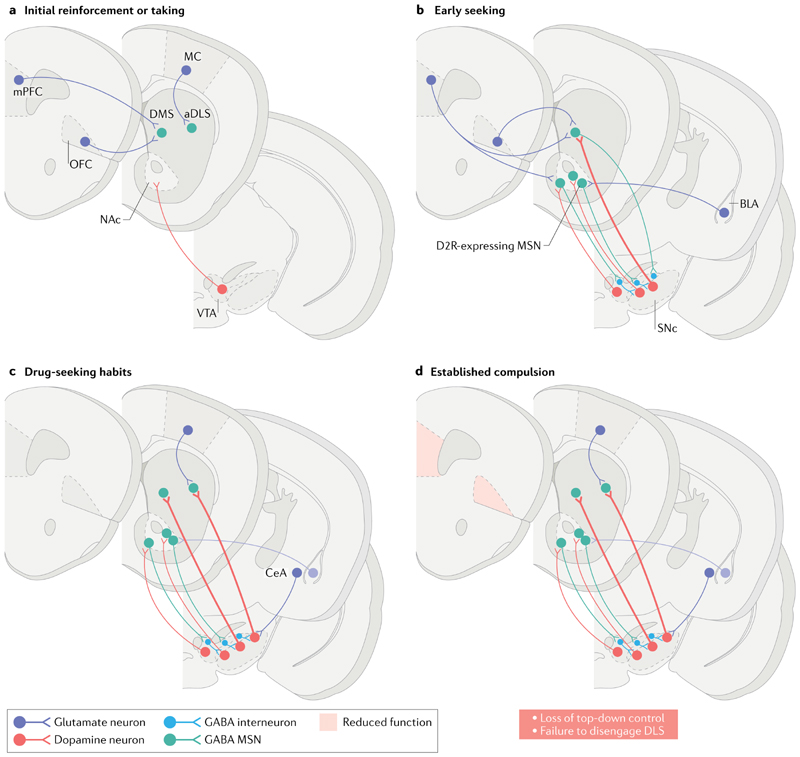
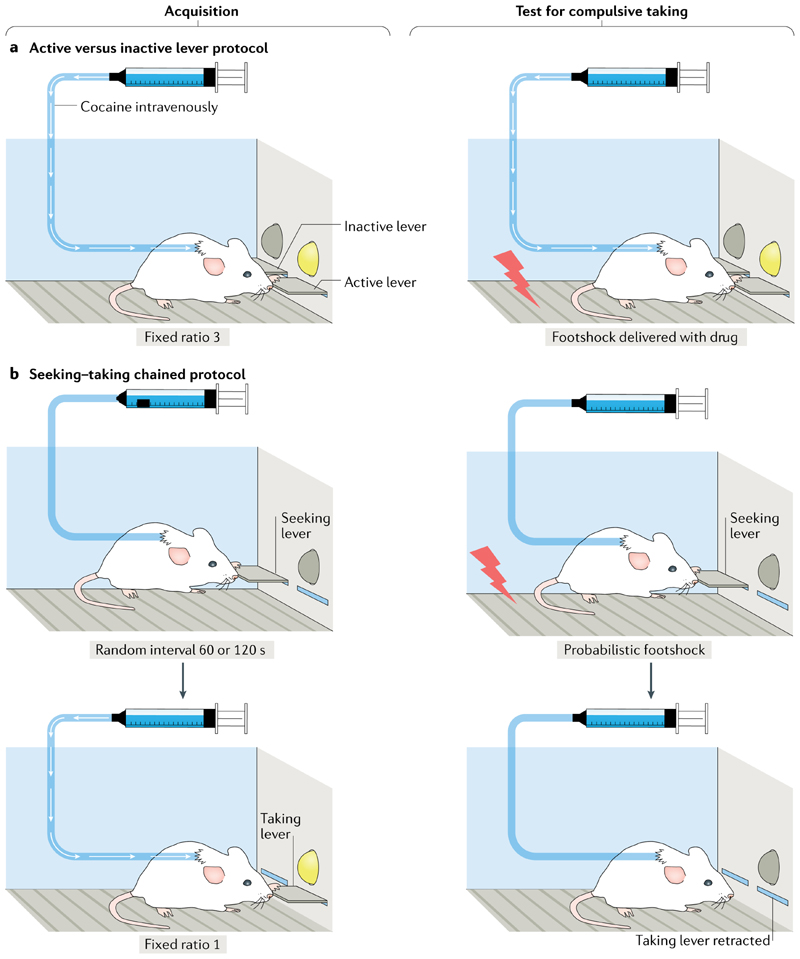
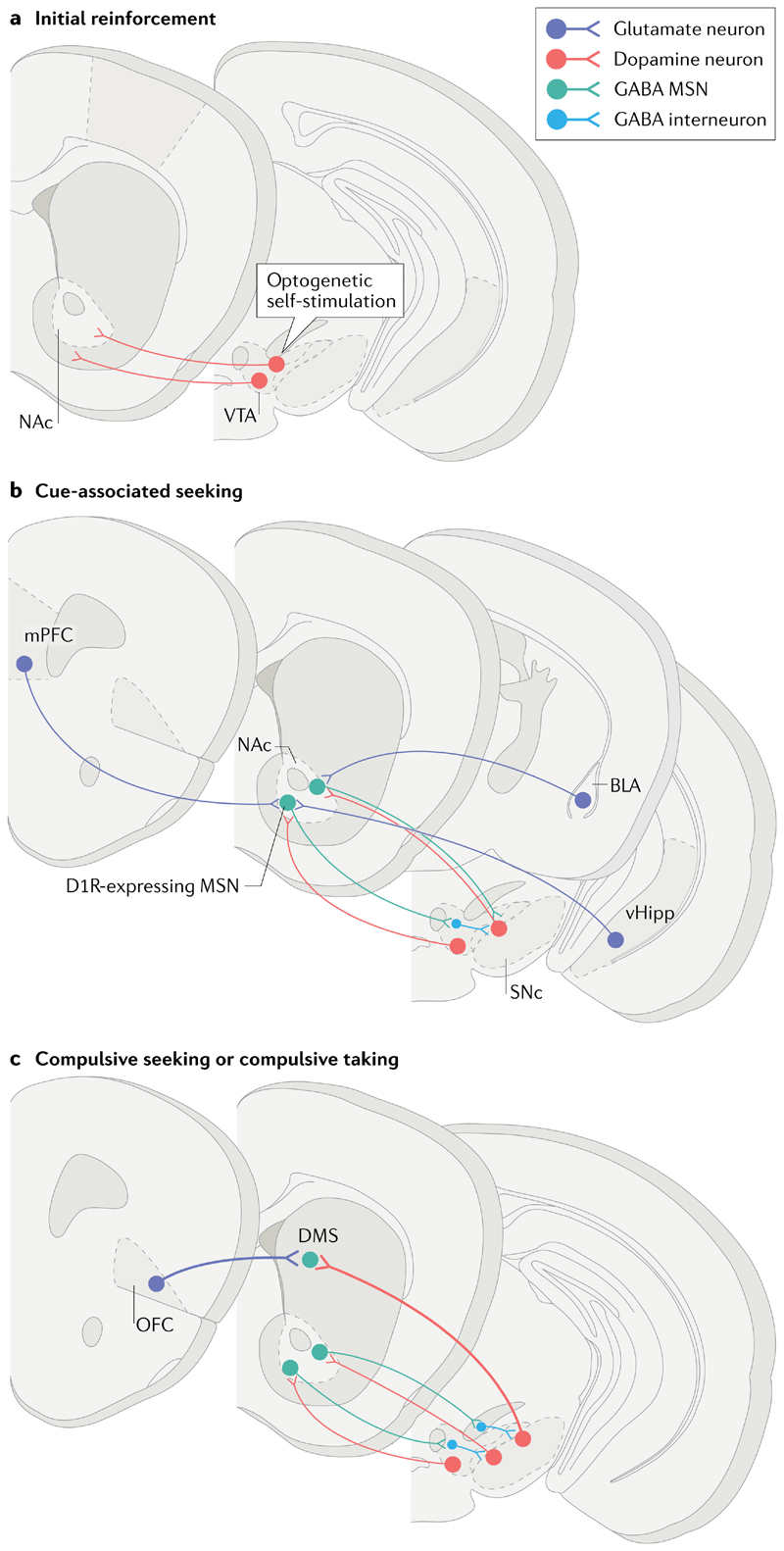
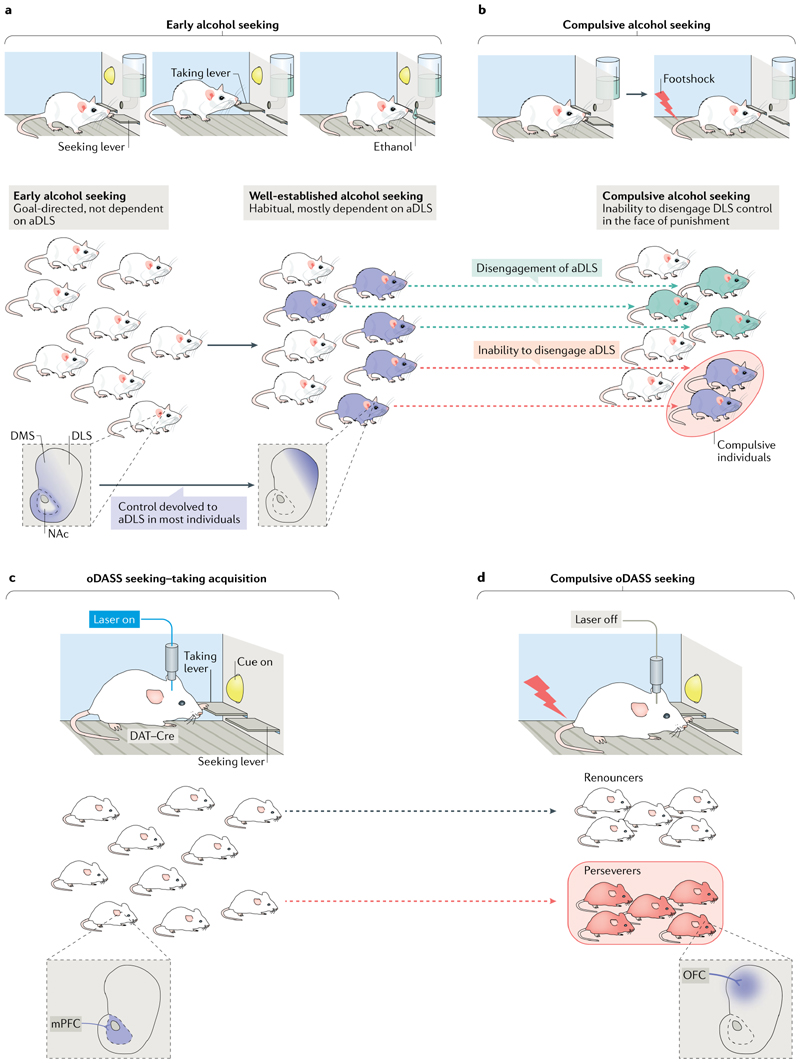
Compulsion is a cardinal symptom of drug addiction (severe substance use disorder). However, compulsion is observed in only a small proportion of individuals who repeatedly seek and use addictive substances. Here, we integrate accounts of the neuropharmacological mechanisms that underlie the transition to compulsion with overarching learning theories, to outline how compulsion develops in addiction. Importantly, we emphasize the conceptual distinctions between compulsive drug-seeking behaviour and compulsive drug-taking behaviour (that is, use). In the latter, an individual cannot stop using a drug despite major negative consequences, possibly reflecting an imbalance in frontostriatal circuits that encode reward and aversion. By contrast, an individual may compulsively seek drugs (that is, persist in seeking drugs despite the negative consequences of doing so) when the neural systems that underlie habitual behaviour dominate goal-directed behavioural systems, and when executive control over this maladaptive behaviour is diminished. This distinction between different aspects of addiction may help to identify its neural substrates and new treatment strategies.
Conflict of interest statement
C.L. has no competing interests. T.W.R. consults for Cambridge Cognition, Unilever, Cassava and Greenfield BioVentures. He holds research grants from Shionogi & Co., Ltd and GlaxoSmithKline plc., and receives royalties from Cambridge Cognition for CANTAB. He also receives editorial honoraria from Springer and Elsevier. B.J.E. has no competing interests.
Figures
References
-
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5. American Psychiatric Association; 2013.
-
- Sanchis-Segura C, Spanagel R. Behavioural assessment of drug reinforcement and addictive features in rodents: an overview. Addict Biol. 2006;11:2–38. - PubMed
