

Light-evoked glutamate transporter EAAT5 activation coordinates with conventional feedback inhibition to control rod bipolar cell output
- PMID: 32233906
- PMCID: PMC7444922
- DOI: 10.1152/jn.00527.2019
Light-evoked glutamate transporter EAAT5 activation coordinates with conventional feedback inhibition to control rod bipolar cell output
Abstract
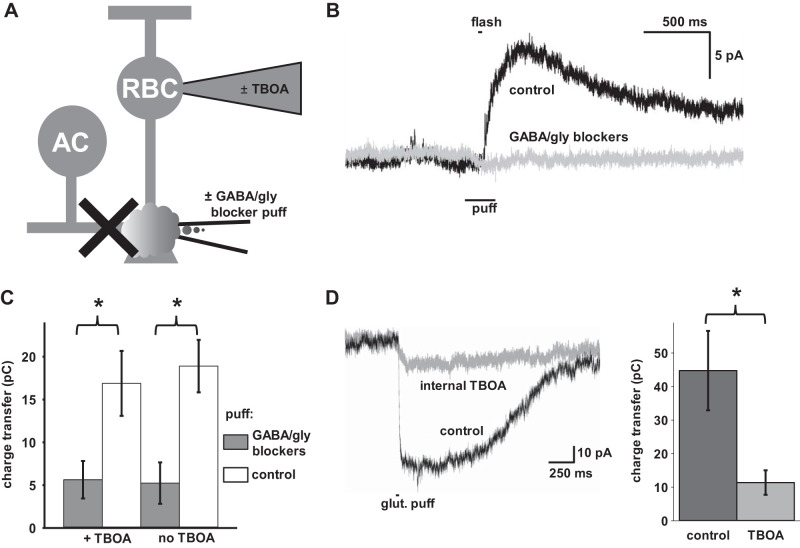
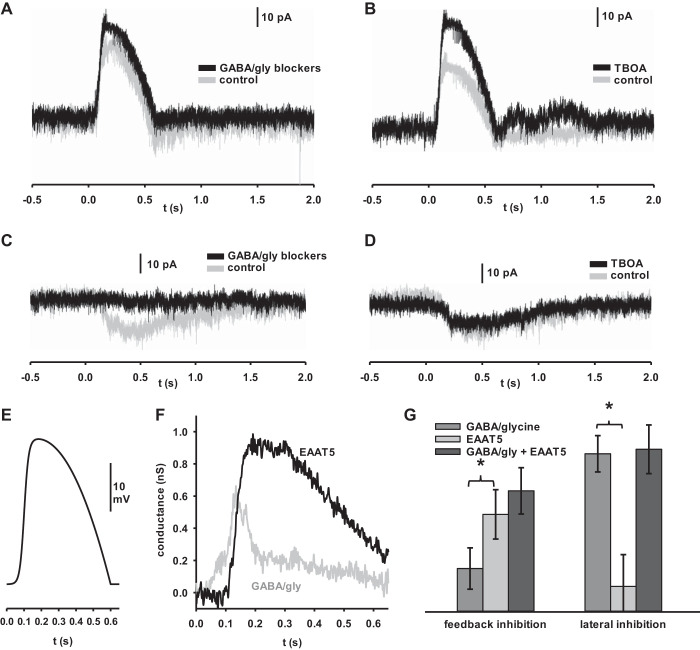
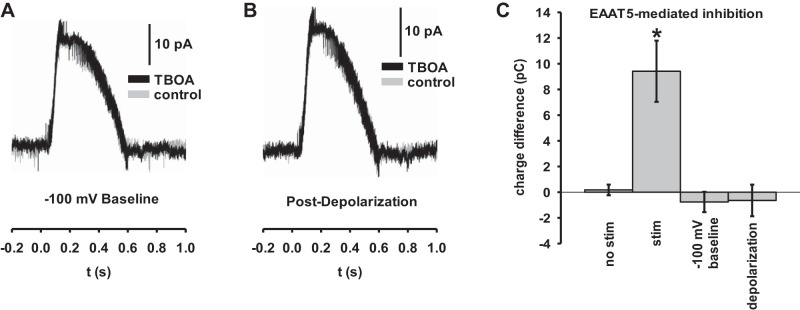
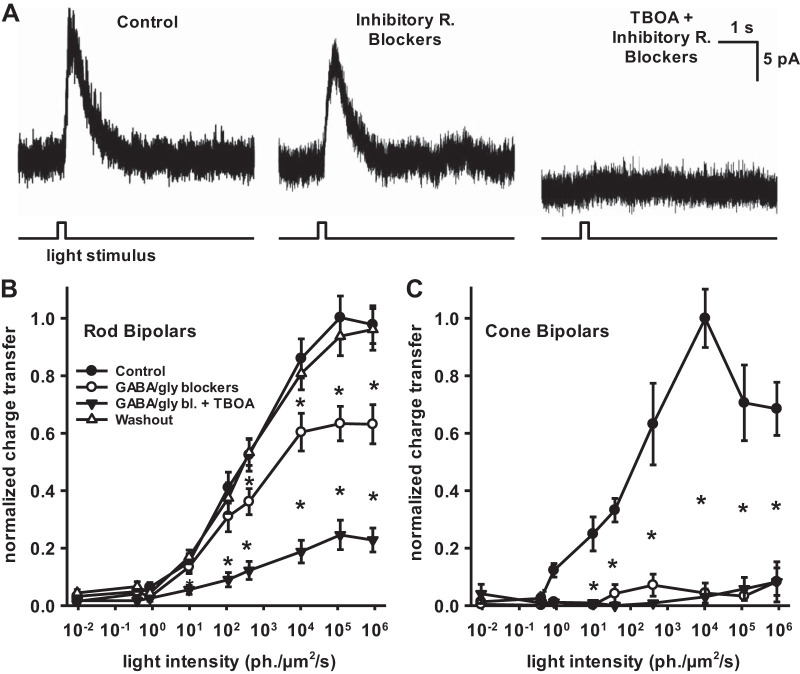
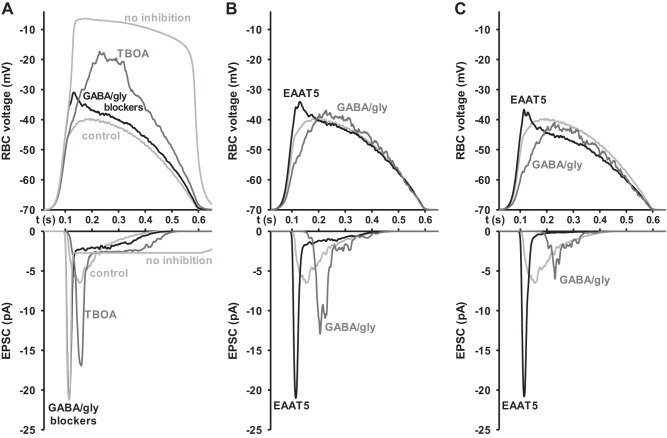
In the retina, modulation of the amplitude of dim visual signals primarily occurs at axon terminals of rod bipolar cells (RBCs). GABA and glycine inhibitory neurotransmitter receptors and the excitatory amino acid transporter 5 (EAAT5) modulate the RBC output. EAATs clear glutamate from the synapse, but they also have a glutamate-gated chloride conductance. EAAT5 acts primarily as an inhibitory glutamate-gated chloride channel. The relative role of visually evoked EAAT5 inhibition compared with GABA and glycine inhibition has not been addressed. In this study, we determine the contribution of EAAT5-mediated inhibition onto RBCs in response to light stimuli in mouse retinal slices. We find differences and similarities in the two forms of inhibition. Our results show that GABA and glycine mediate nearly all lateral inhibition onto RBCs, as EAAT5 is solely a mediator of RBC feedback inhibition. We also find that EAAT5 and conventional GABA inhibition both contribute to feedback inhibition at all stimulus intensities. Finally, our in silico modeling compares and contrasts EAAT5-mediated to GABA- and glycine-mediated feedback inhibition. Both forms of inhibition have a substantial impact on synaptic transmission to the postsynaptic AII amacrine cell. Our results suggest that the late phase EAAT5 inhibition acts with the early phase conventional, reciprocal GABA inhibition to modulate the rod signaling pathway between rod bipolar cells and their downstream synaptic targets.NEW & NOTEWORTHY Excitatory amino acid transporter 5 (EAAT5) glutamate transporters have a chloride channel that is strongly activated by glutamate, which modulates excitatory signaling. We found that EAAT5 is a major contributor to feedback inhibition on rod bipolar cells. Inhibition to rod bipolar cells is also mediated by GABA and glycine. GABA and glycine mediate the early phase of feedback inhibition, and EAAT5 mediates a more delayed inhibition. Together, inhibitory transmitters and EAAT5 coordinate to mediate feedback inhibition, controlling neuronal output.
Keywords: EAAT5; glucose transporter; retina; rod bipolar cell.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases