

Pericyte abnormalities precede strial capillary basement membrane thickening in Alport mice
- PMID: 32234583
- PMCID: PMC7491280
- DOI: 10.1016/j.heares.2020.107935
Pericyte abnormalities precede strial capillary basement membrane thickening in Alport mice
Abstract
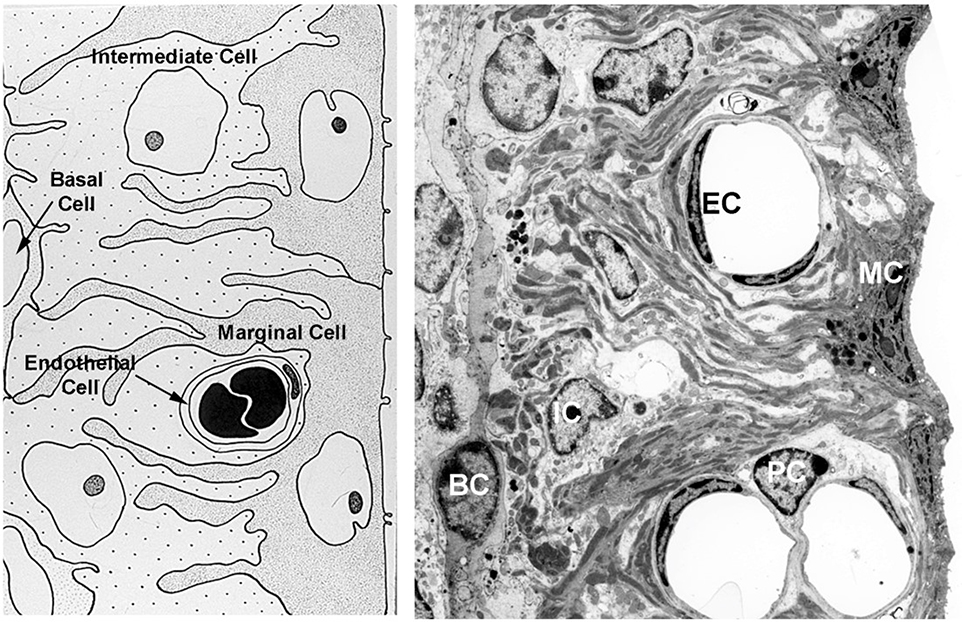
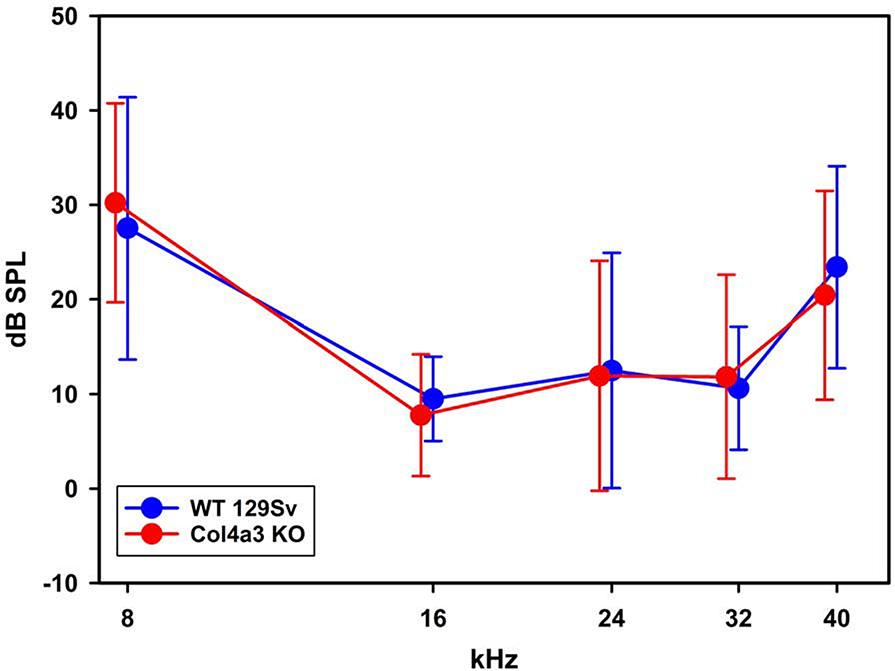
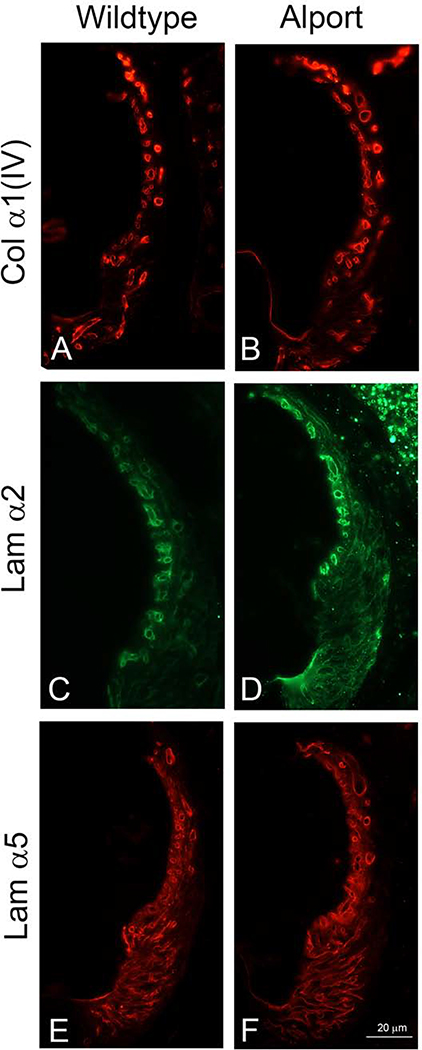
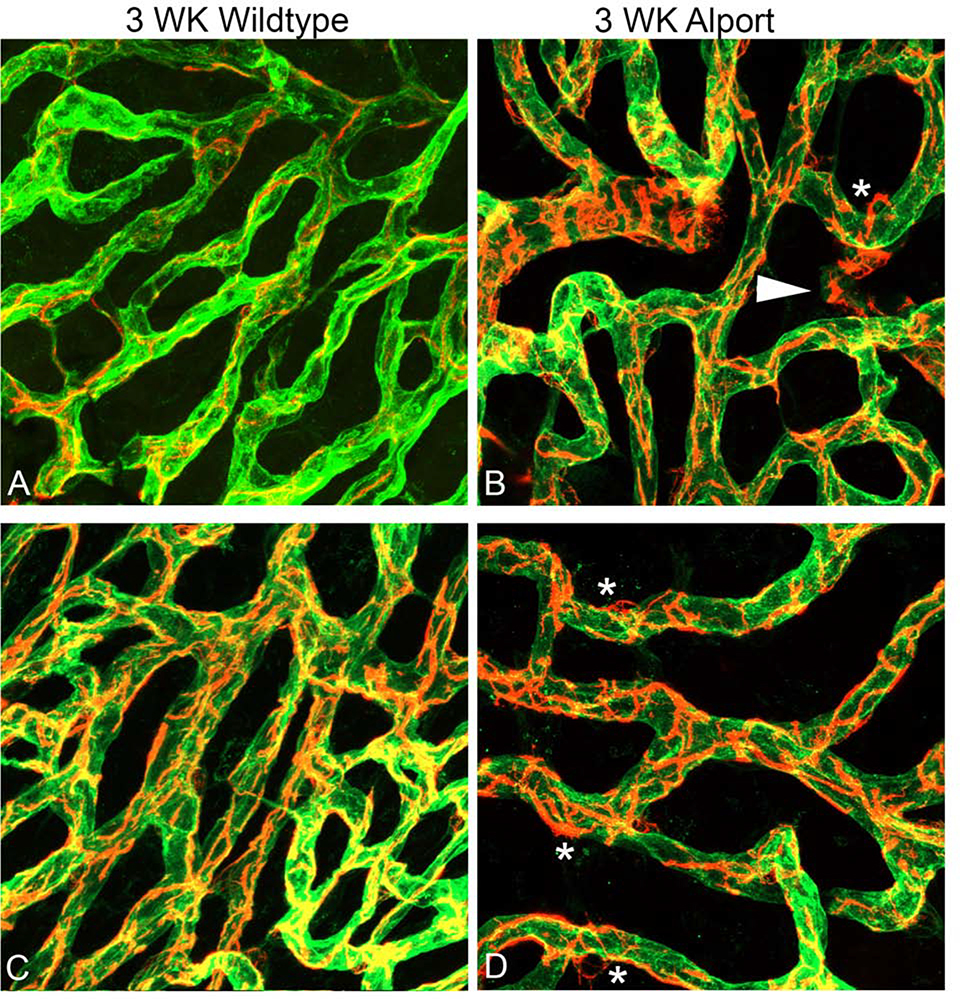
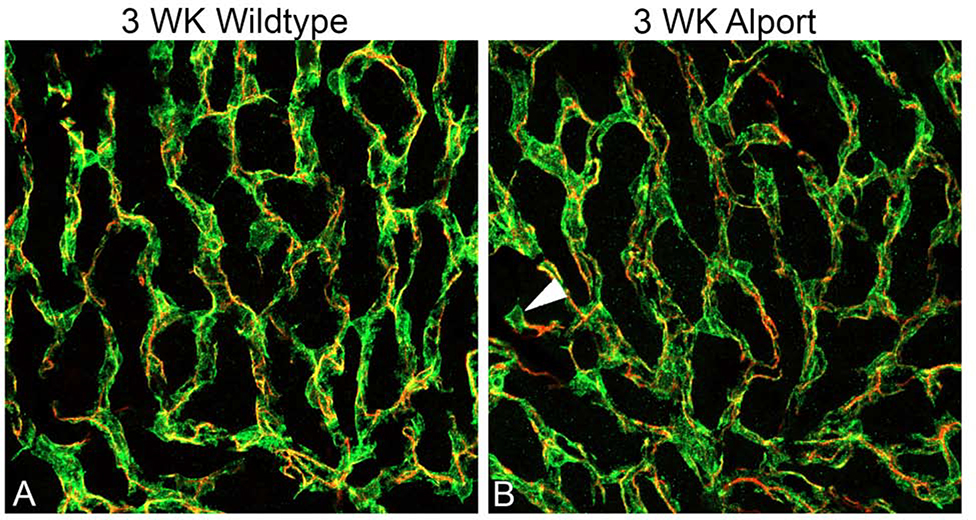
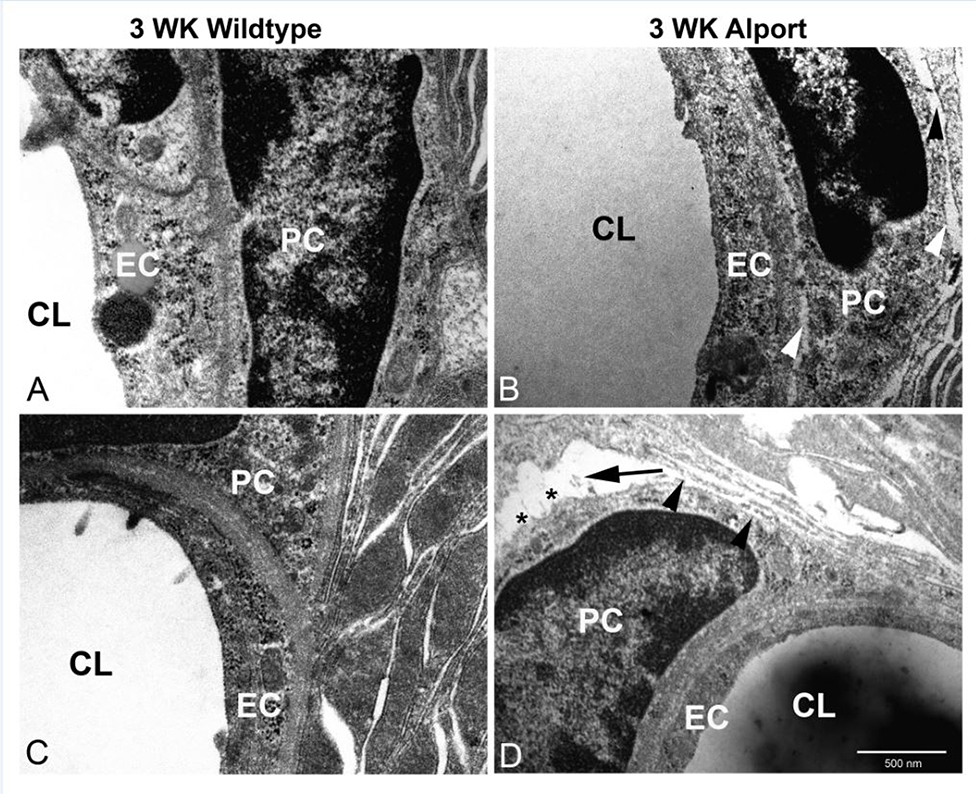
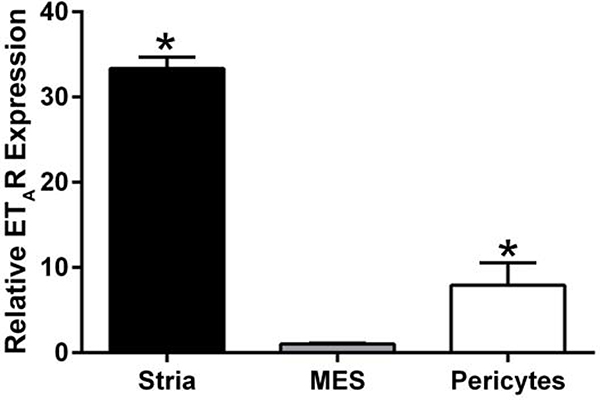
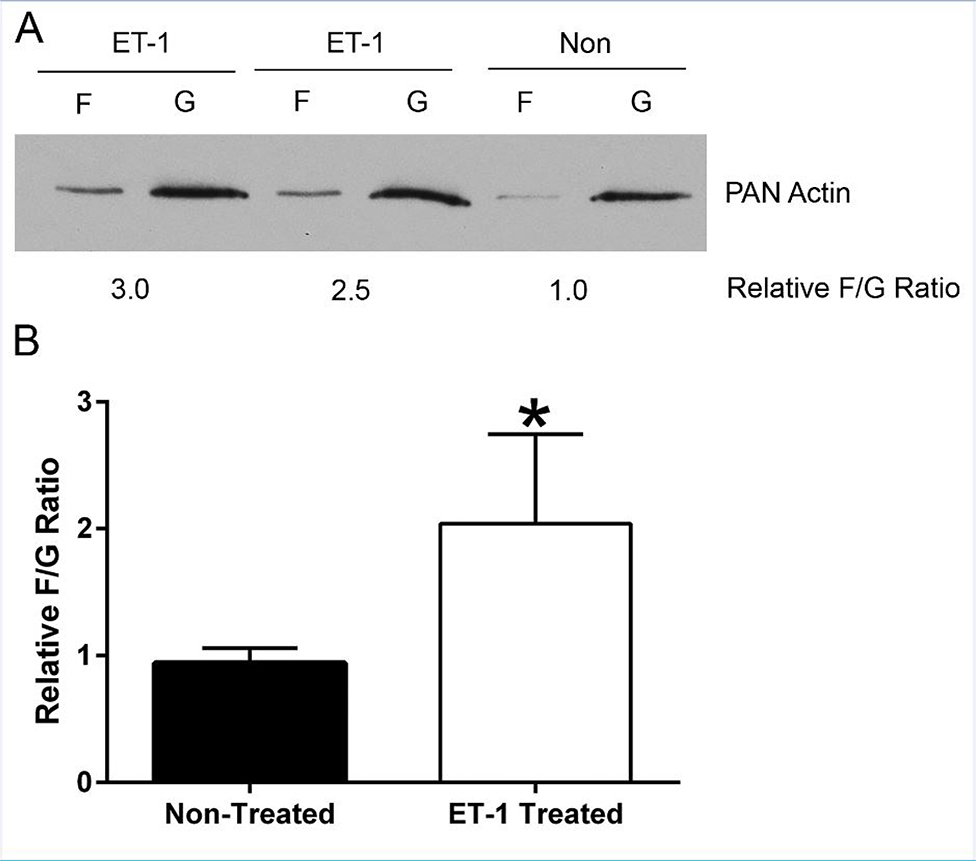
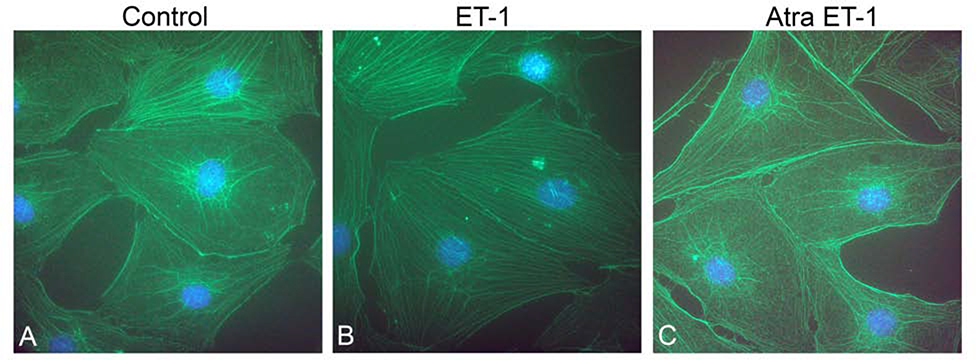
In 129 Sv autosomal Alport mice, the strial capillary basement membranes (SCBMs) progressively thicken between 5 and 9 weeks of age resulting in a hypoxic microenvironment with metabolic stress and induction of pro-inflammatory cytokines and chemokines. These events occur concomitant with a drop in endocochlear potential and a susceptibility to noise-induced hearing loss under conditions that do not permanently affect age/strain-matched littermates. Here we aimed to gain an understanding of events that occur before the onset of SCBM thickening. Alport stria has normal thickness and shows levels of extracellular matrix (ECM) molecules in the SCBMs commensurate with wild-type mice. Hearing thresholds in the 3-week Alport mice do not differ from those of wild-type mice. We performed RNAseq analysis using RNA from stria vascularis isolated from 3-week Alport mice and wild type littermates. Data was processed using Ingenuity Pathway Analysis software and further distilled using manual procedures. RNAseq analysis revealed significant dysregulation of genes involved in cell adhesion, cell migration, formation of protrusions, and both actin and tubulin cytoskeletal dynamics. Overall, the data suggested changes in the cellular architecture of the stria might be apparent. To test this notion, we performed dual immunofluorescence analysis on whole mounts of the stria vascularis from these same animals stained with anti-isolectin gs-ib4 (endothelial cell marker) and anti-desmin (pericyte marker) antibodies. The results showed evidence of pericyte detachment and migration as well as the formation of membrane ruffling on pericytes in z-stacked confocal images from Alport mice compared to wild type littermates. This was confirmed by TEM analysis. Earlier work from our lab showed that endothelin A receptor blockade prevents SCBM thickening and ECM accumulation in the SCBMs. Treating cultured pericytes with endothelin-1 induced actin cytoskeletal rearrangement, increasing the ratio of filamentous to globular actin. Collectively, these findings suggest that the change in type IV collagen composition in the Alport SCBMs results in cellular insult to the pericyte compartment, activating detachment and altered cytoskeletal dynamics. These events precede SCBM thickening and hearing loss in Alport mice, and thus constitute the earliest event so far recognized in Alport strial pathology.
Keywords: Alport syndrome; Pericyte; RNA-seq; Strial capillary basement membrane.
Copyright © 2020 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors of this work have no conflicts of interest to disclose.
Figures

References
-
- Bertlich M, Ihler F, Weiss BG, Freytag S, Strupp M, Canis M 2017. Cochlear pericytes are capable of decreasing capillary diameter in vivo after tumor necrosis factor exposure. Otol. Neurotol 38, e545–e550. - PubMed
-
- Cosgrove D, Meehan DT, Grunkemeyer JA, Kornak JM, Sayers R, Hunter WT Samuelson, G.C. 1996. Collagen 4A3 knockout: A mouse model for autosomal Alport Syndrome. Genes and Development 10, 2981–2992. - PubMed
-
- Cosgrove DE, Samuelson G, Meehan DT, Miller C, McGee J, Walsh EJ & Siegel M 1998. Ultrastructural, physiological, and molecular defects in the inner ear of a gene-knockout mouse model for autosomal Alport syndrome. Hear. Res 121, 84–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
