

Indian Hedgehog regulates senescence in bone marrow-derived mesenchymal stem cell through modulation of ROS/mTOR/4EBP1, p70S6K1/2 pathway
- PMID: 32235006
- PMCID: PMC7185126
- DOI: 10.18632/aging.102958
Indian Hedgehog regulates senescence in bone marrow-derived mesenchymal stem cell through modulation of ROS/mTOR/4EBP1, p70S6K1/2 pathway
Abstract
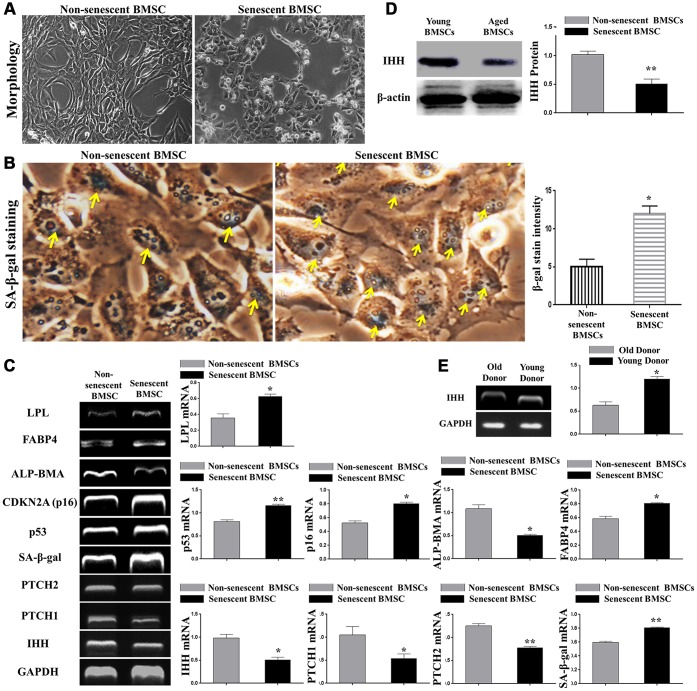
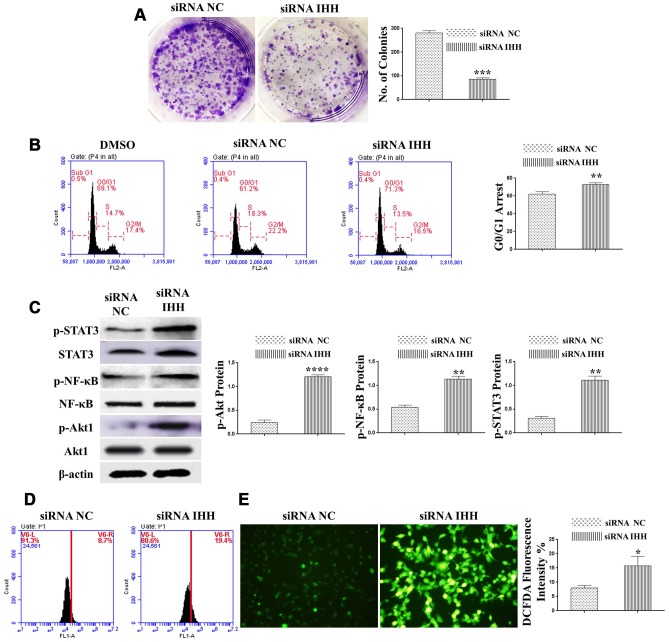
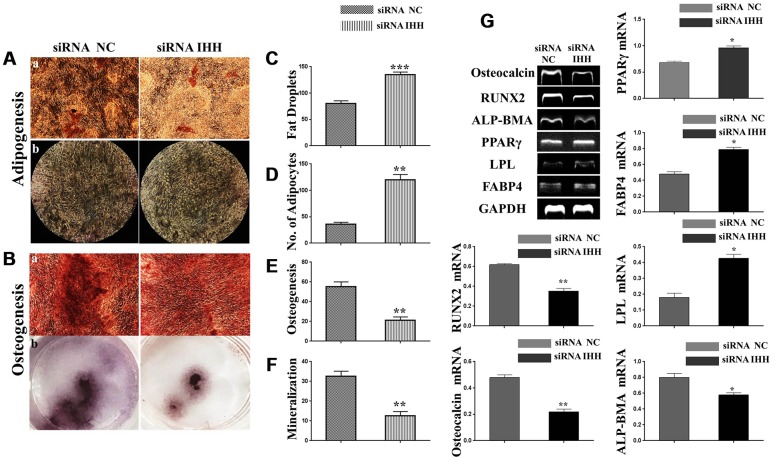
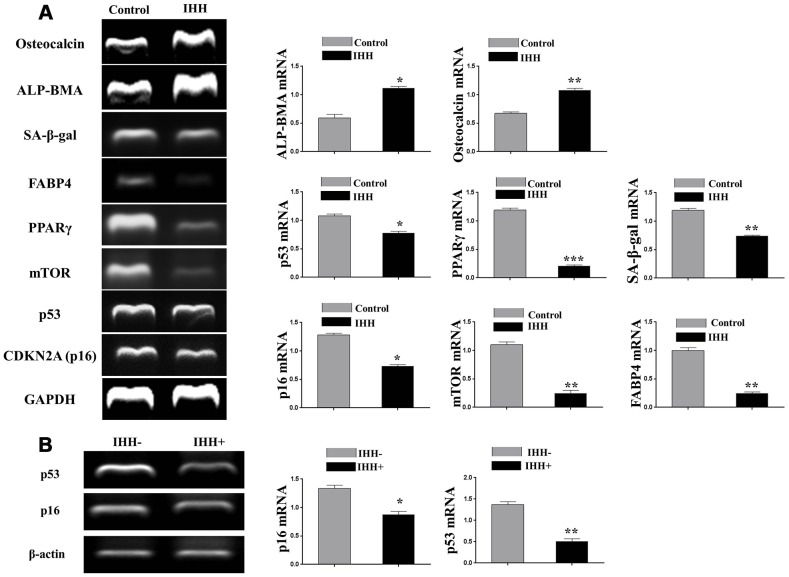
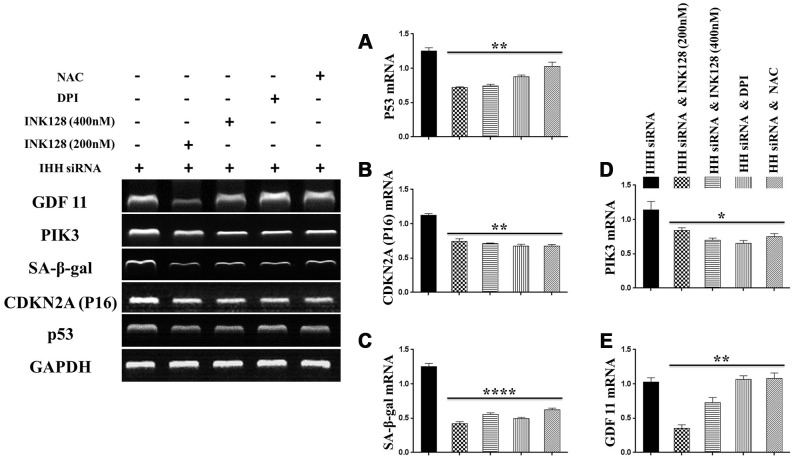
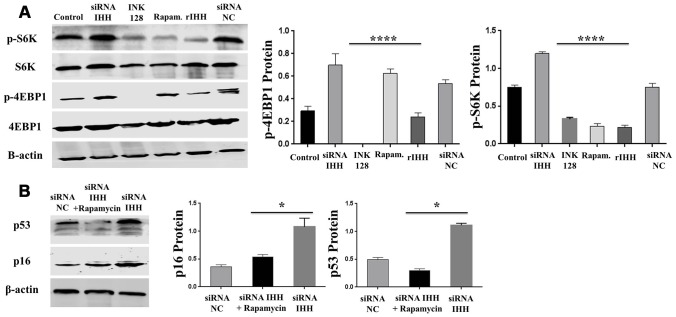
Premature senescence of bone marrow-derived mesenchymal stem cells (BMSC) remains a major concern for their application clinically. Hedgehog signaling has been reported to regulate aging-associated markers and MSC skewed differentiation. Indian Hedgehog (IHH) is a ligand of Hedgehog intracellular pathway considered as an inducer in chondrogenesis of human BMSC. However, the role of IHH in the aging of BMSC is still unclear. This study explored the role IHH in the senescence of BMSC obtained from human samples and senescent mice. Isolated BMSC were transfected with IHH siRNA or incubated with exogenous IHH protein and the mechanisms of aging and differentiation investigated. Moreover, the interactions between IHH, and mammalian target of rapamycin (mTOR) and reactive oxygen species (ROS) were evaluated using the corresponding inhibitors and antioxidants. BMSC transfected with IHH siRNA showed characteristics of senescence-associated features including increased senescence-associated β-galactosidase activity (SA-β-gal), induction of cell cycle inhibitors (p53/p16), development of senescence-associated secretory phenotype (SASP), activation of ROS and mTOR pathways as well as the promotion of skewed differentiation. Interestingly, BMSC treatment with IHH protein reversed the senescence markers and corrected biased differentiation. Moreover, IHH shortage-induced senescence signs were compromised after mTOR and ROS inhibition. Our findings presented anti-aging activity for IHH in BMSC through down-regulation of ROS/mTOR pathways. This discovery might contribute to increasing the therapeutic, immunomodulatory and regenerative potency of BMSC and introduce a novel remedy in the management of aging-related diseases.
Keywords: Indian hedgehog; aging; differentiation; mammalian target of rapamycin; mesenchymal stem cell.
Conflict of interest statement
Figures




References
-
- Duscher D, Rennert RC, Januszyk M, Anghel E, Maan ZN, Whittam AJ, Perez MG, Kosaraju R, Hu MS, Walmsley GG, Atashroo D, Khong S, Butte AJ, Gurtner GC. Aging disrupts cell subpopulation dynamics and diminishes the function of mesenchymal stem cells. Sci Rep. 2014; 4:7144. 10.1038/srep07144 - DOI - PMC - PubMed
-
- Bustos ML, Huleihel L, Kapetanaki MG, Lino-Cardenas CL, Mroz L, Ellis BM, McVerry BJ, Richards TJ, Kaminski N, Cerdenes N, Mora AL, Rojas M. Aging mesenchymal stem cells fail to protect because of impaired migration and antiinflammatory response. Am J Respir Crit Care Med. 2014; 189:787–98. 10.1164/rccm.201306-1043OC - DOI - PMC - PubMed
-
- Yukata K, Xie C, Li TF, Takahata M, Hoak D, Kondabolu S, Zhang X, Awad HA, Schwarz EM, Beck CA, Jonason JH, O’Keefe RJ. Aging periosteal progenitor cells have reduced regenerative responsiveness to bone injury and to the anabolic actions of PTH 1-34 treatment. Bone. 2014; 62:79–89. 10.1016/j.bone.2014.02.002 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
