

Role of hyaluronan in regulating self-renewal and osteogenic differentiation of mesenchymal stromal cells and pre-osteoblasts
- PMID: 32236725
- PMCID: PMC7544712
- DOI: 10.1007/s00784-020-03259-8
Role of hyaluronan in regulating self-renewal and osteogenic differentiation of mesenchymal stromal cells and pre-osteoblasts
Abstract
Objectives: The aim of the study was to investigate the impact of two hyaluronan (HA) formulations on the osteogenic potential of osteoblast precursors.
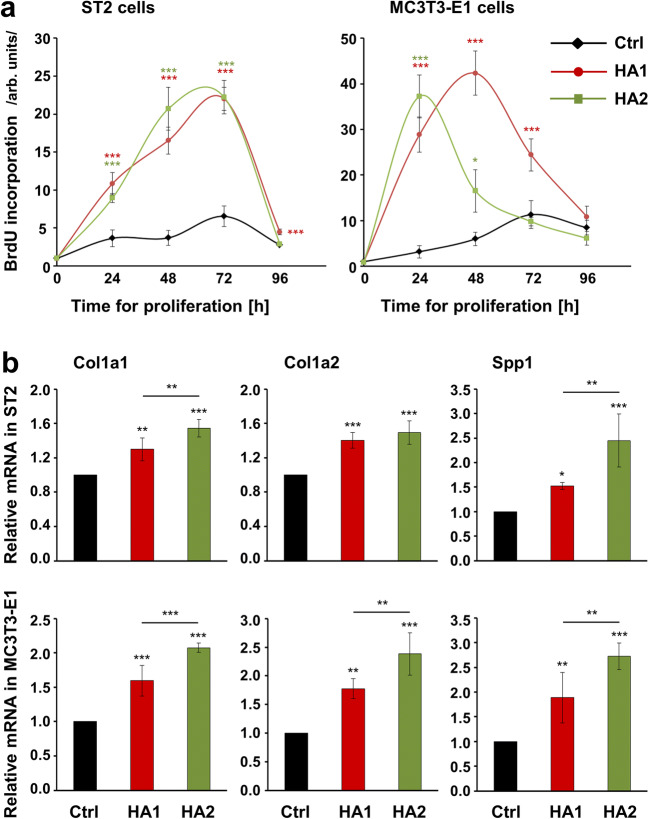
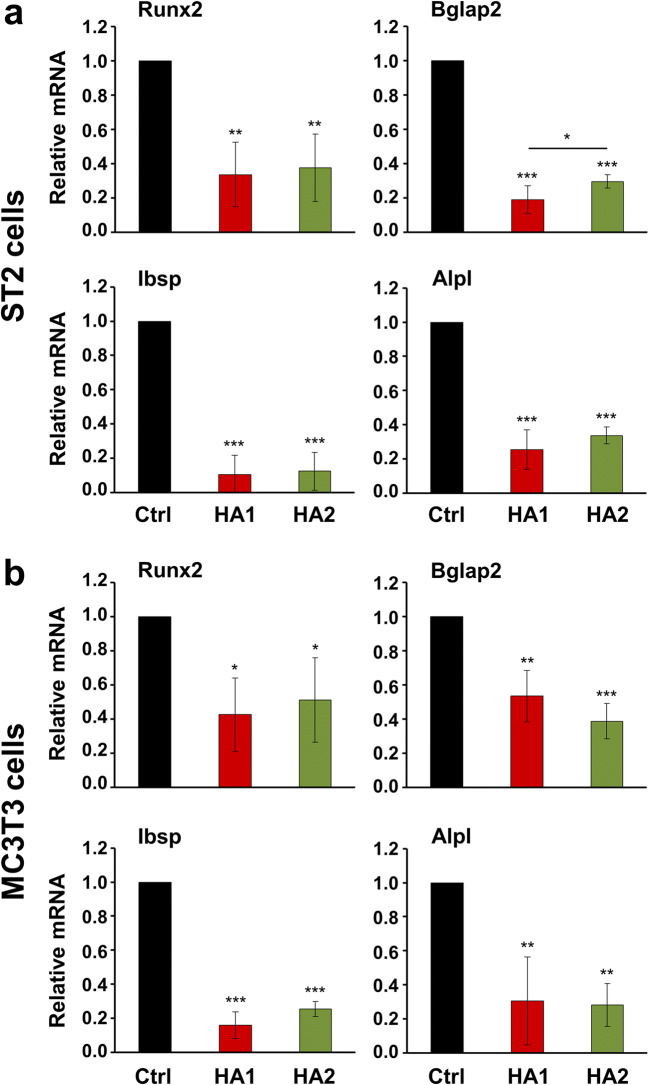
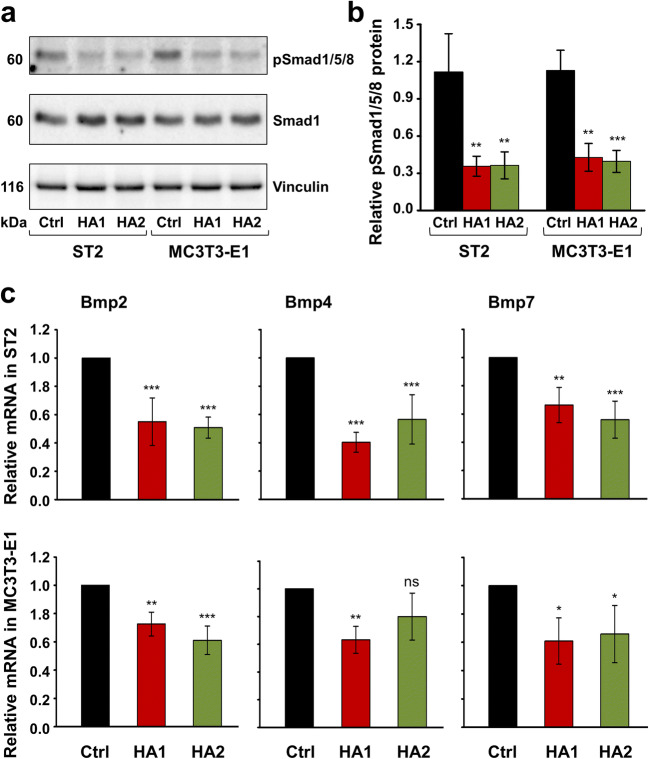
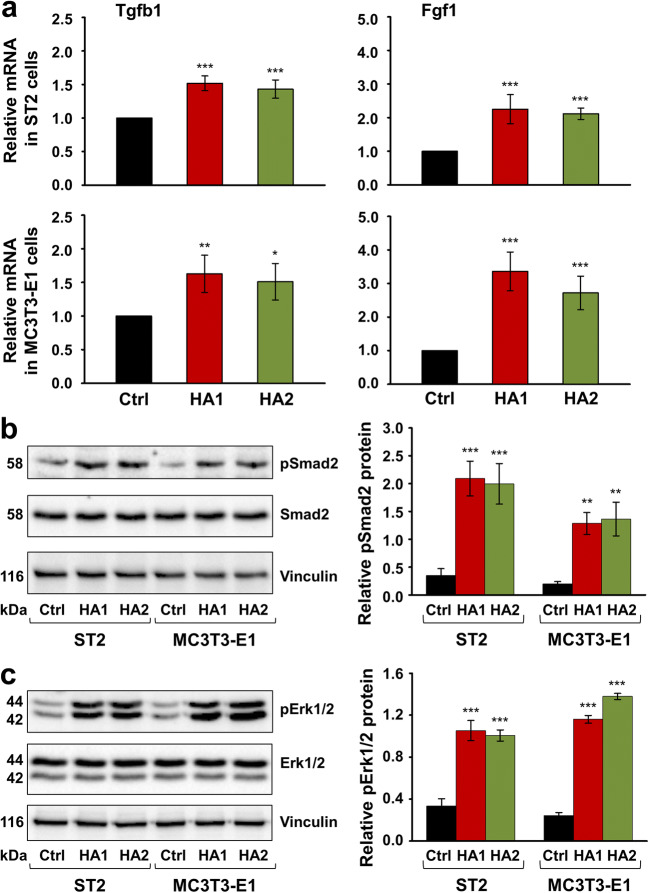
Materials and methods: Proliferation rates of HA-treated mesenchymal stromal ST2 and pre-osteoblastic MC3T3-E1 cells were determined by 5-bromo-20-deoxyuridine (BrdU) assay. Expression of genes encoding osteogenic differentiation markers, critical growth, and stemness factors as well as activation of downstream signaling pathways in the HA-treated cells were analyzed by quantitative reverse transcription-polymerase chain reaction (qRT-PCR) and immunoblot techniques.
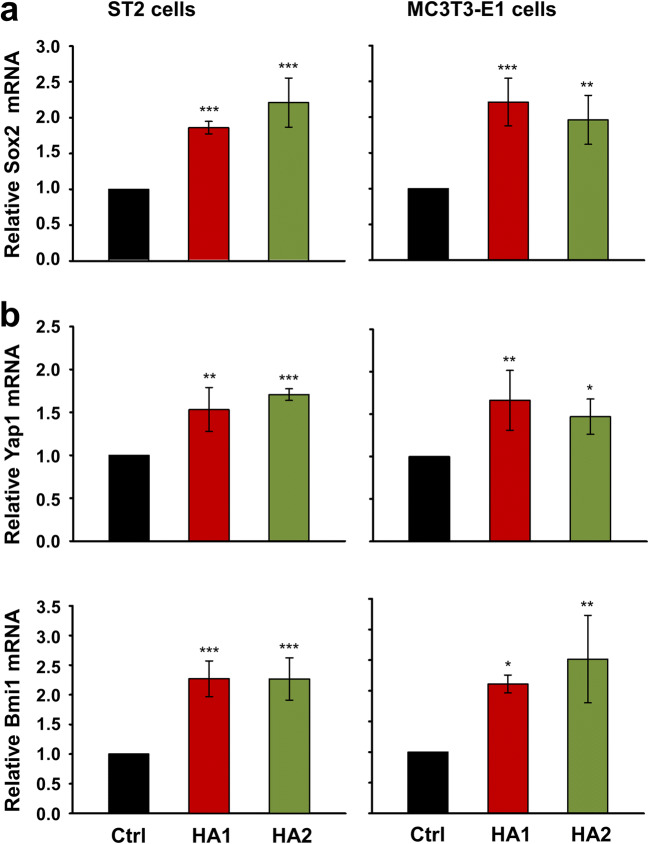
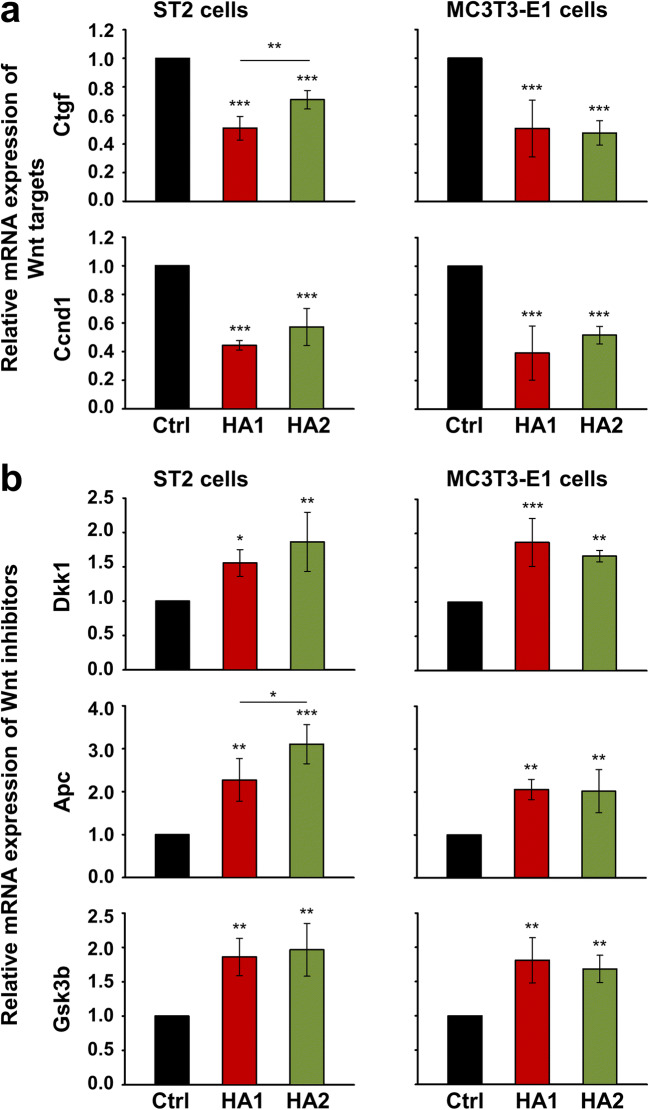
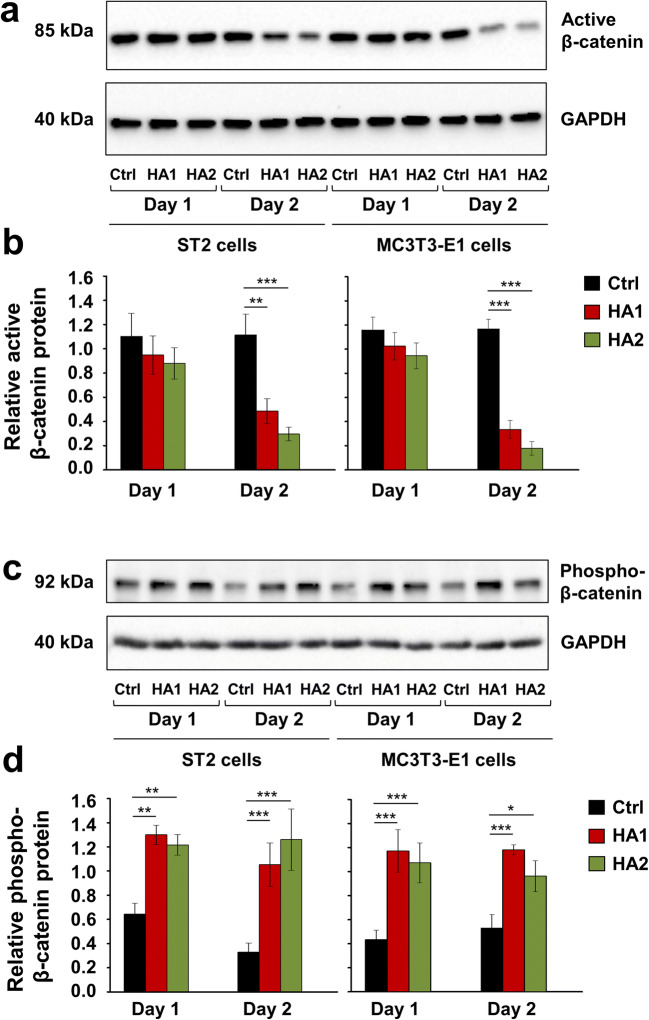
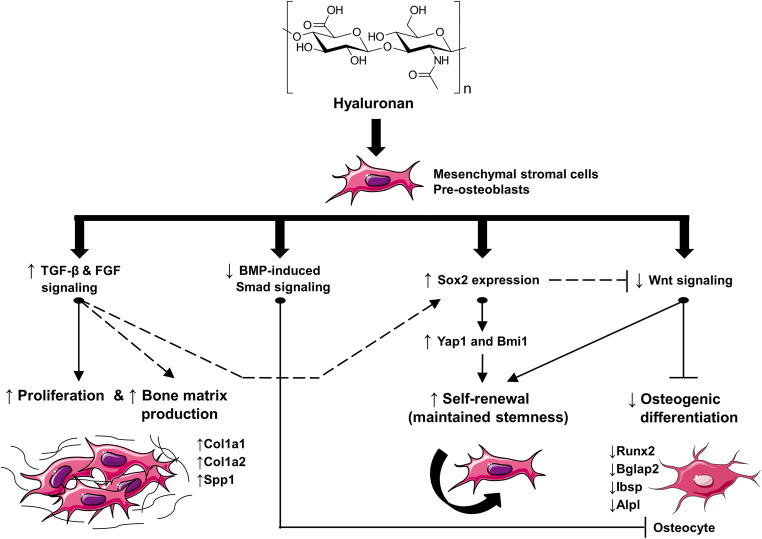
Results: The investigated HAs strongly stimulated the growth of the osteoprogenitor lines and enhanced the expression of genes encoding bone matrix proteins. However, expression of late osteogenic differentiation markers was significantly inhibited, accompanied by decreased bone morphogenetic protein (BMP) signaling. The expression of genes encoding transforming growth factor-β1 (TGF-β1) and fibroblast growth factor-1 (FGF-1) as well as the phosphorylation of the downstream signaling molecules Smad2 and Erk1/2 were enhanced upon HA treatment. We observed significant upregulation of the transcription factor Sox2 and its direct transcription targets and critical stemness genes, Yap1 and Bmi1, in HA-treated cells. Moreover, prominent targets of the canonical Wnt signaling pathway showed reduced expression, whereas inhibitors of the pathway were considerably upregulated. We detected decrease of active β-catenin levels in HA-treated cells due to β-catenin being phosphorylated and, thus, targeted for degradation.
Conclusions: HA strongly induces the growth of osteoprogenitors and maintains their stemness, thus potentially regulating the balance between self-renewal and differentiation during bone regeneration following reconstructive oral surgeries.
Clinical relevance: Addition of HA to deficient bone or bony defects during implant or reconstructive periodontal surgeries may be a viable approach for expanding adult stem cells without losing their replicative and differentiation capabilities.
Keywords: Bone and soft tissue regeneration; Extracellular matrix; Gene expression; Growth factors; Hyaluronic acid; Stemness.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
