

Identification of Reptarenaviruses, Hartmaniviruses, and a Novel Chuvirus in Captive Native Brazilian Boa Constrictors with Boid Inclusion Body Disease
- PMID: 32238580
- PMCID: PMC7269426
- DOI: 10.1128/JVI.00001-20
Identification of Reptarenaviruses, Hartmaniviruses, and a Novel Chuvirus in Captive Native Brazilian Boa Constrictors with Boid Inclusion Body Disease
Abstract
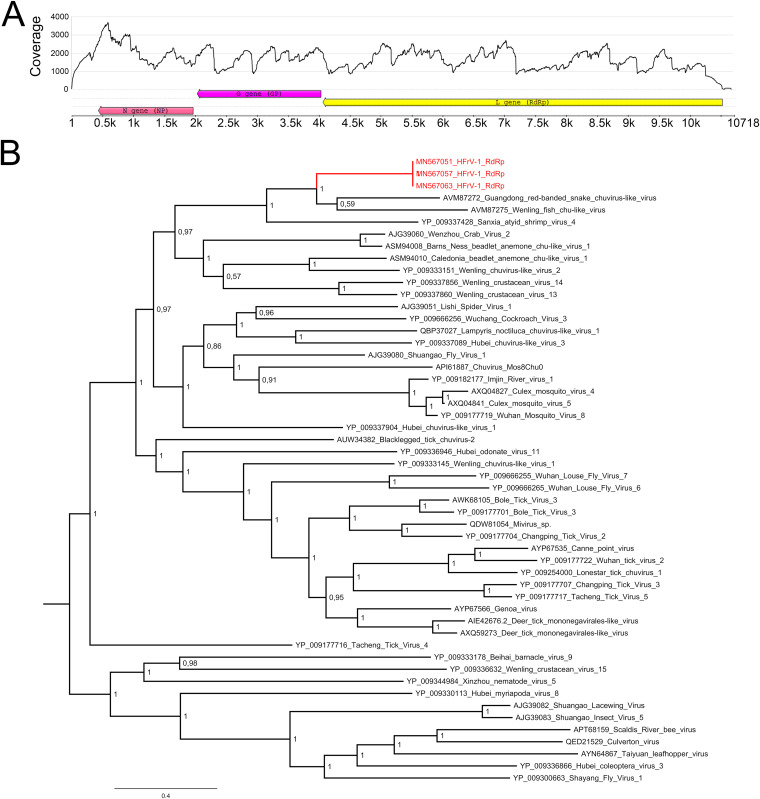
Boid inclusion body disease (BIBD) is a transmissible viral disease of captive snakes that causes severe losses in snake collections worldwide. It is caused by reptarenavirus infection, which can persist over several years without overt signs but is generally associated with the eventual death of the affected snakes. Thus far, reports have confirmed the existence of reptarenaviruses in captive snakes in North America, Europe, Asia, and Australia, but there is no evidence that it also occurs in wild snakes. BIBD affects boa species within the subfamily Boinae and pythons in the family Pythonidae, the habitats of which do not naturally overlap. Here, we studied Brazilian captive snakes with BIBD using a metatranscriptomic approach, and we report the identification of novel reptarenaviruses, hartmaniviruses, and a new species in the family Chuviridae The reptarenavirus L segments identified are divergent enough to represent six novel species, while we found only a single novel reptarenavirus S segment. Until now, hartmaniviruses had been identified only in European captive boas with BIBD, and the present results increase the number of known hartmaniviruses from four to six. The newly identified chuvirus showed 38.4%, 40.9%, and 48.1% amino acid identity to the nucleoprotein, glycoprotein, and RNA-dependent RNA polymerase, respectively, of its closest relative, Guangdong red-banded snake chuvirus-like virus. Although we cannot rule out the possibility that the found viruses originated from imported snakes, the results suggest that the viruses could circulate in indigenous snake populations.IMPORTANCE Boid inclusion body disease (BIBD), caused by reptarenavirus infection, affects captive snake populations worldwide, but the reservoir hosts of reptarenaviruses remain unknown. Here, we report the identification of novel reptarenaviruses, hartmaniviruses, and a chuvirus in captive Brazilian boas with BIBD. Three of the four snakes studied showed coinfection with all three viruses, and one of the snakes harbored three novel reptarenavirus L segments and one novel S segment. The samples originated from collections with Brazilian indigenous snakes only, which could indicate that these viruses circulate in wild snakes. The findings could further indicate that boid snakes are the natural reservoir of reptarena- and hartmaniviruses commonly found in captive snakes. The snakes infected with the novel chuvirus all suffered from BIBD; it is therefore not possible to comment on its potential pathogenicity and contribution to the observed changes in the present case material.
Keywords: arenavirus; boa constrictor; boid inclusion body disease; chuvirus; hartmanivirus; reptarenavirus.
Copyright © 2020 American Society for Microbiology.
Figures
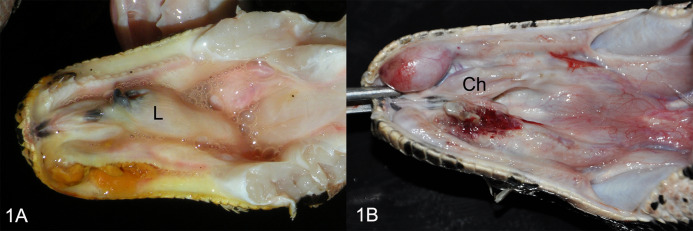
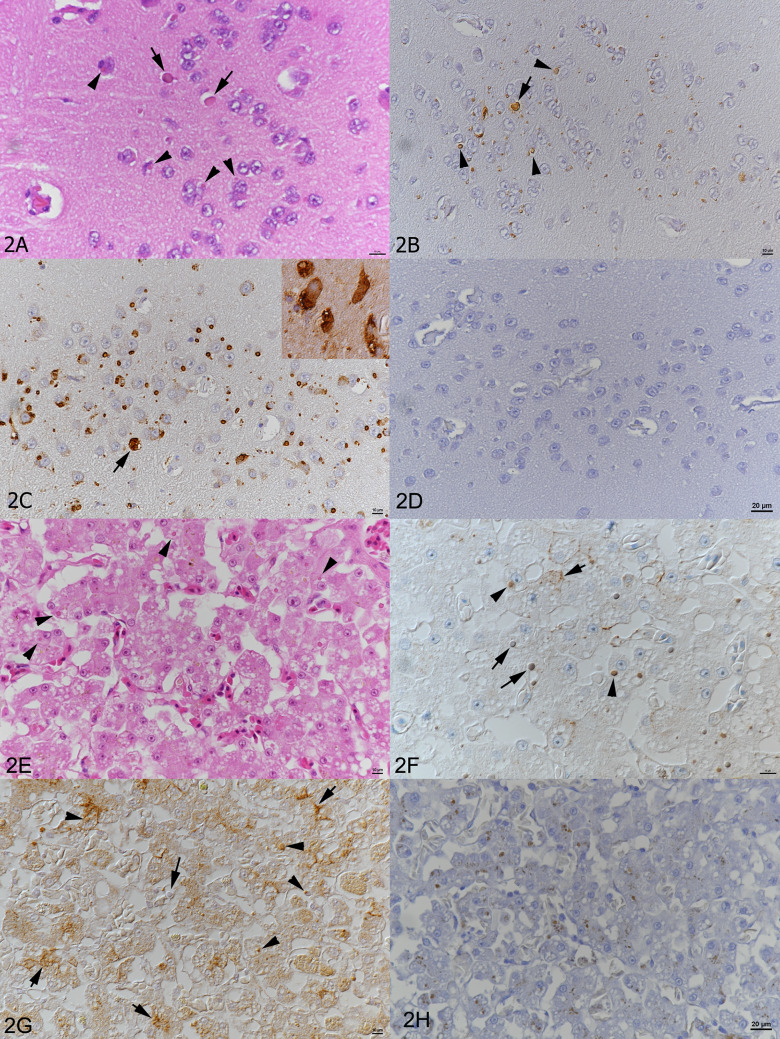
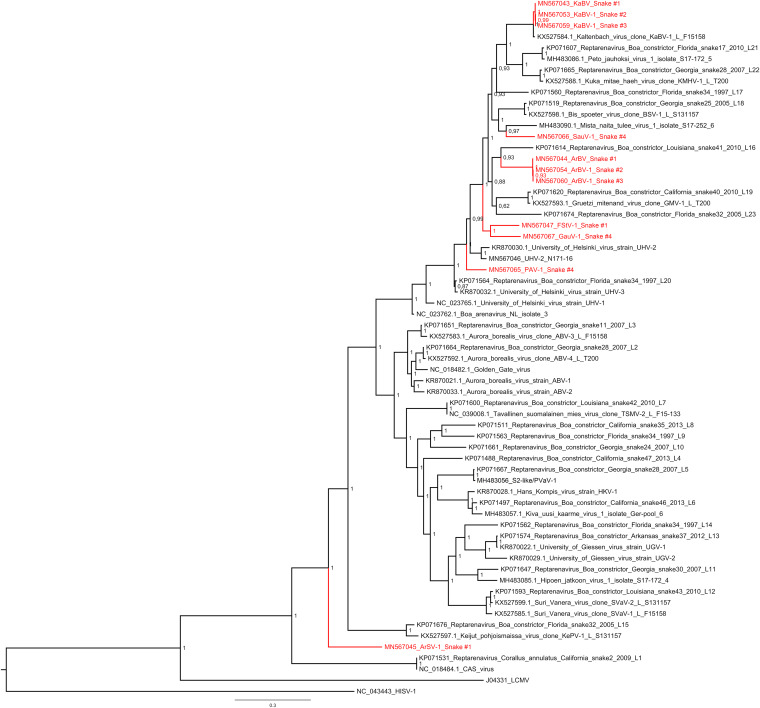
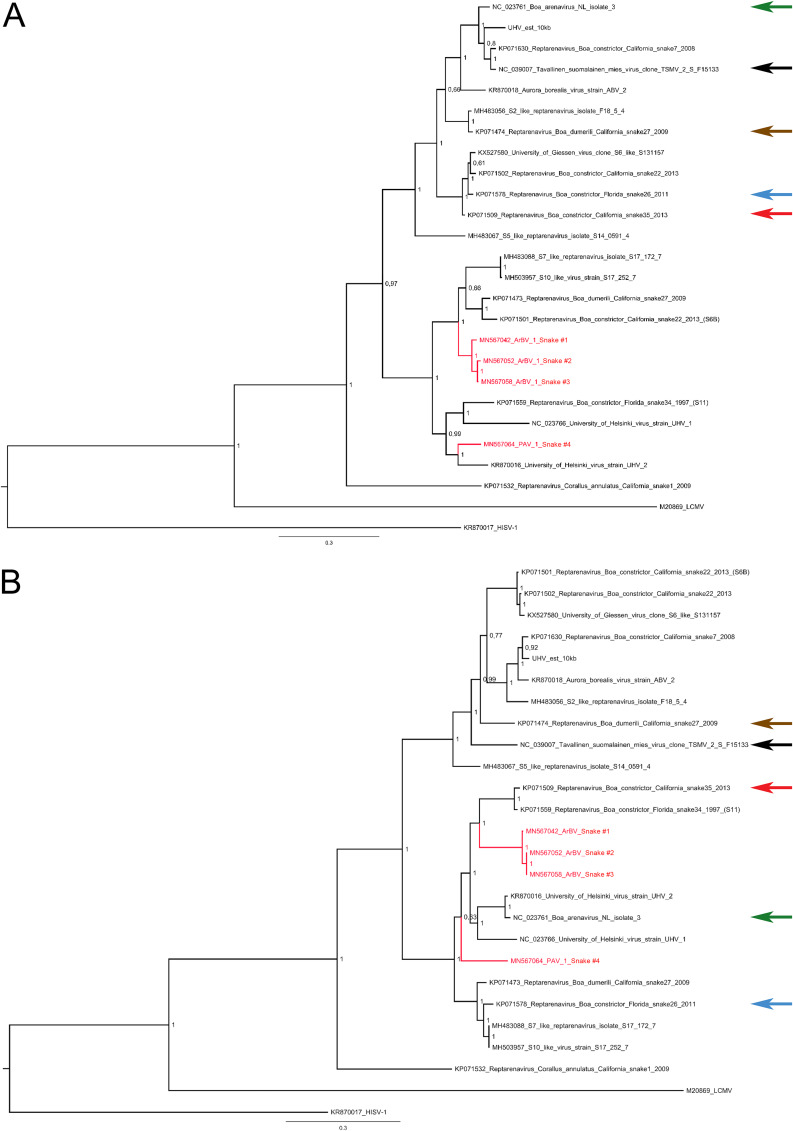

References
-
- Chang L, Jacobson ER. 2010. Inclusion body disease, a worldwide infectious disease of boid snakes: a review. J Exotic Pet Med 19:216–225. doi: 10.1053/j.jepm.2010.07.014. - DOI
-
- Schumacher J, Jacobson ER, Homer BL, Gaskin JM. 1994. Inclusion body disease in boid snakes. J Zoo Wildl Med 25:511–524.
-
- Stenglein MD, Sanders C, Kistler AL, Ruby JG, Franco JY, Reavill DR, Dunker F, Derisi JL. 2012. Identification, characterization, and in vitro culture of highly divergent arenaviruses from boa constrictors and annulated tree boas: candidate etiological agents for snake inclusion body disease. mBio 3:e00180-12. doi: 10.1128/mBio.00180-12. - DOI - PMC - PubMed
-
- Hetzel U, Sironen T, Laurinmaki P, Liljeroos L, Patjas A, Henttonen H, Vaheri A, Artelt A, Kipar A, Butcher SJ, Vapalahti O, Hepojoki J. 2013. Isolation, identification, and characterization of novel arenaviruses, the etiological agents of boid inclusion body disease. J Virol 87:10918–10935. doi: 10.1128/JVI.01123-13. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
