

Bilateral and unilateral odor processing and odor perception
- PMID: 32238904
- PMCID: PMC7113286
- DOI: 10.1038/s42003-020-0876-6
Bilateral and unilateral odor processing and odor perception
Abstract
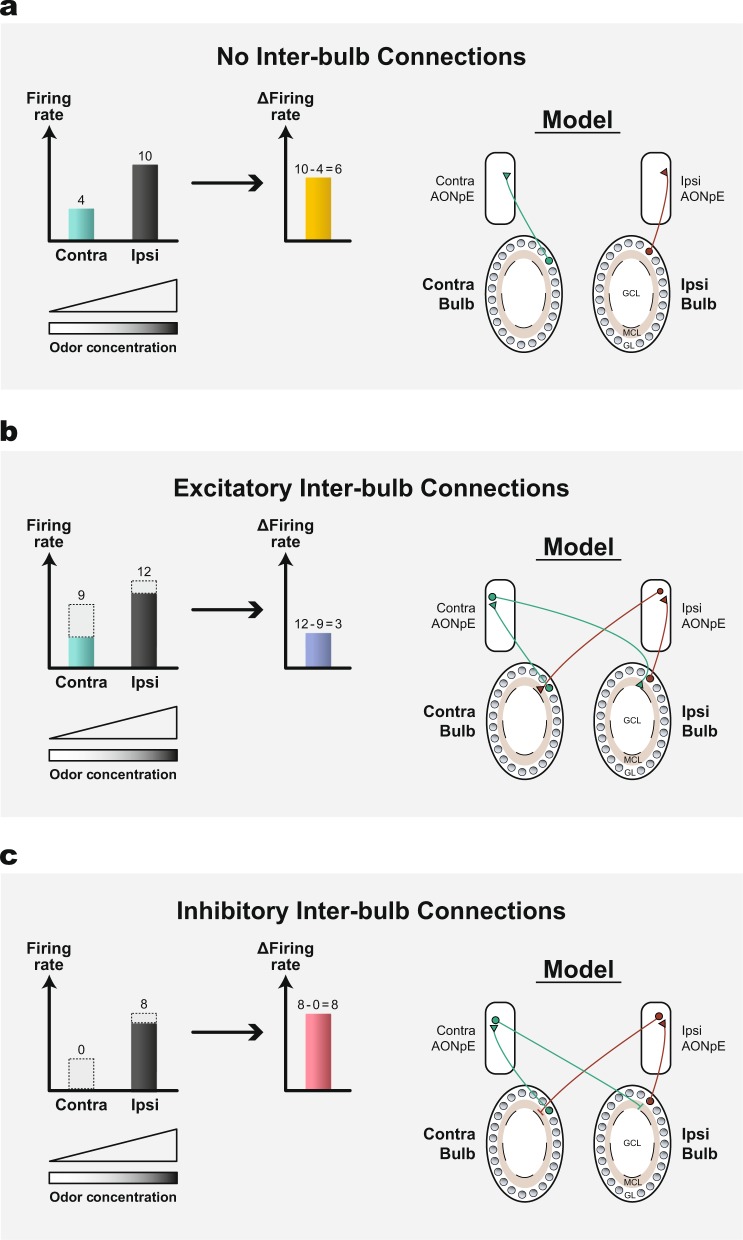
Imagine smelling a novel perfume with only one nostril and then smelling it again with the other nostril. Clearly, you can tell that it is the same perfume both times. This simple experiment demonstrates that odor information is shared across both hemispheres to enable perceptual unity. In many sensory systems, perceptual unity is believed to be mediated by inter-hemispheric connections between iso-functional cortical regions. However, in the olfactory system, the underlying neural mechanisms that enable this coordination are unclear because the two olfactory cortices are not topographically organized and do not seem to have homotypic inter-hemispheric mapping. This review presents recent advances in determining which aspects of odor information are processed unilaterally or bilaterally, and how odor information is shared across the two hemispheres. We argue that understanding the mechanisms of inter-hemispheric coordination can provide valuable insights that are hard to achieve when focusing on one hemisphere alone.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Bilateral Alignment of Receptive Fields in the Olfactory Cortex.eNeuro. 2024 Nov 6;11(11):ENEURO.0155-24.2024. doi: 10.1523/ENEURO.0155-24.2024. Print 2024 Nov. eNeuro. 2024. PMID: 39433407 Free PMC article.
-
A Mirror-Symmetric Excitatory Link Coordinates Odor Maps across Olfactory Bulbs and Enables Odor Perceptual Unity.Neuron. 2018 Aug 22;99(4):800-813.e6. doi: 10.1016/j.neuron.2018.07.012. Epub 2018 Aug 2. Neuron. 2018. PMID: 30078580
-
The olfactory bulb: coding and processing of odor molecule information.Science. 1999 Oct 22;286(5440):711-5. doi: 10.1126/science.286.5440.711. Science. 1999. PMID: 10531048 Review.
-
Editorial for the special issue "Olfactory Coding and Circuitries".Cell Tissue Res. 2021 Jan;383(1):1-6. doi: 10.1007/s00441-020-03389-1. Cell Tissue Res. 2021. PMID: 33447882 Free PMC article. No abstract available.
-
Function follows form: ecological constraints on odor codes and olfactory percepts.Curr Opin Neurobiol. 2009 Aug;19(4):422-9. doi: 10.1016/j.conb.2009.07.012. Epub 2009 Aug 9. Curr Opin Neurobiol. 2009. PMID: 19671493 Free PMC article. Review.
Cited by
-
An optofluidic platform for interrogating chemosensory behavior and brainwide neural representation in larval zebrafish.Nat Commun. 2023 Jan 14;14(1):227. doi: 10.1038/s41467-023-35836-2. Nat Commun. 2023. PMID: 36641479 Free PMC article.
-
Generating parallel representations of position and identity in the olfactory system.Cell. 2023 Jun 8;186(12):2556-2573.e22. doi: 10.1016/j.cell.2023.04.038. Epub 2023 May 25. Cell. 2023. PMID: 37236194 Free PMC article.
-
Organizational Principles of the Centrifugal Projections to the Olfactory Bulb.Int J Mol Sci. 2023 Feb 26;24(5):4579. doi: 10.3390/ijms24054579. Int J Mol Sci. 2023. PMID: 36902010 Free PMC article.
-
Bilateral Alignment of Receptive Fields in the Olfactory Cortex.eNeuro. 2024 Nov 6;11(11):ENEURO.0155-24.2024. doi: 10.1523/ENEURO.0155-24.2024. Print 2024 Nov. eNeuro. 2024. PMID: 39433407 Free PMC article.
-
One nose but two nostrils: Learn to align with sparse connections between two olfactory cortices.ArXiv [Preprint]. 2024 May 6:arXiv:2405.03602v1. ArXiv. 2024. PMID: 38764587 Free PMC article. Preprint.
References
-
- Boycott, B. B., J. D. Organization of the primate retina: Light microscopy, with an appendix: a second type of midget bipolar cell in the primate retina. Philos. Trans. R. Soc. Lond. B Biol. Sci. 255, 109–184 (1969).
-
- Shepherd, G. M. The Synaptic Organization of the Brain (Oxford University Press, 2004).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
