

Replication-inducible vaccinia virus vectors with enhanced safety in vivo
- PMID: 32240193
- PMCID: PMC7117657
- DOI: 10.1371/journal.pone.0230711
Replication-inducible vaccinia virus vectors with enhanced safety in vivo
Abstract
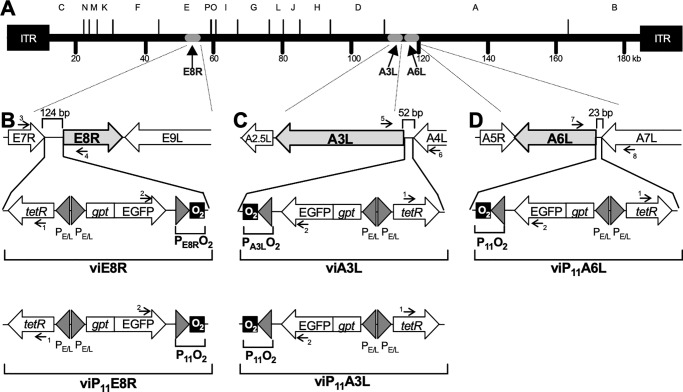
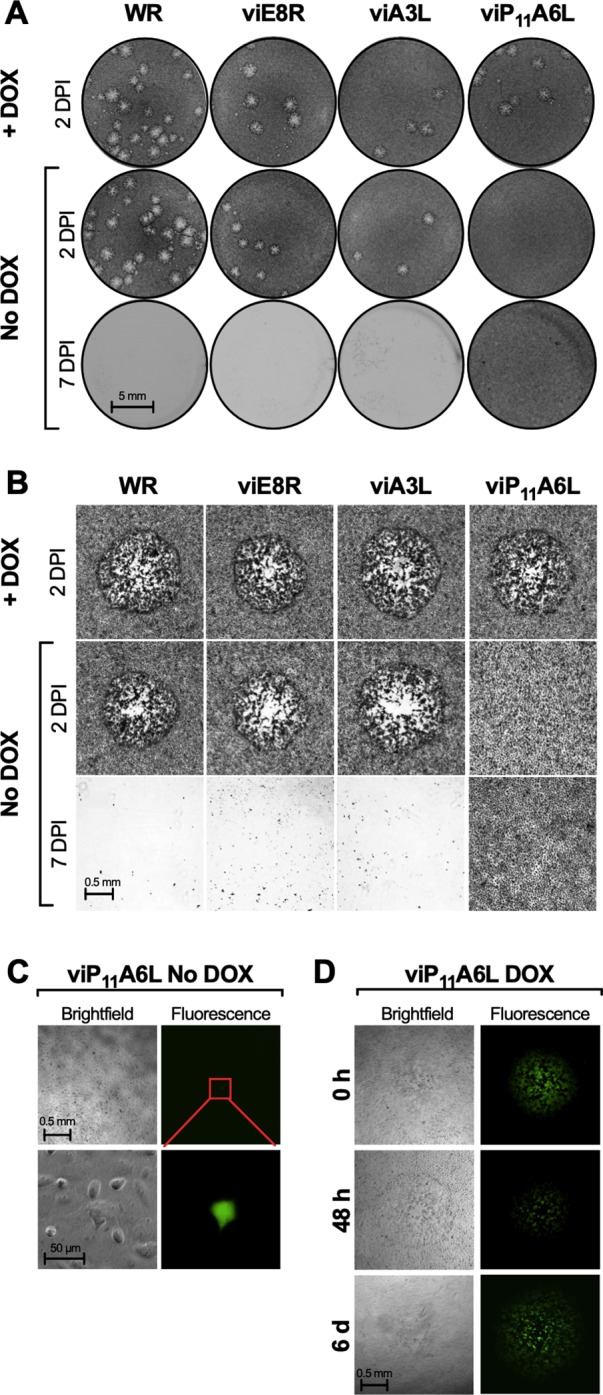
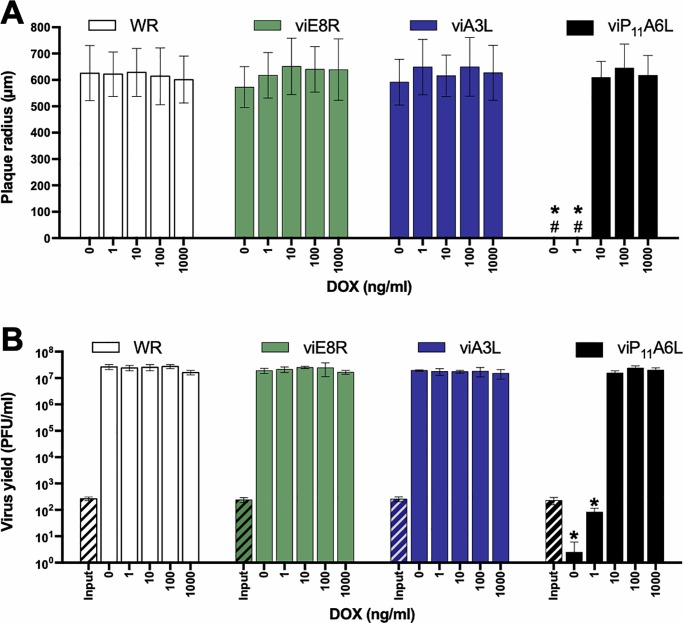
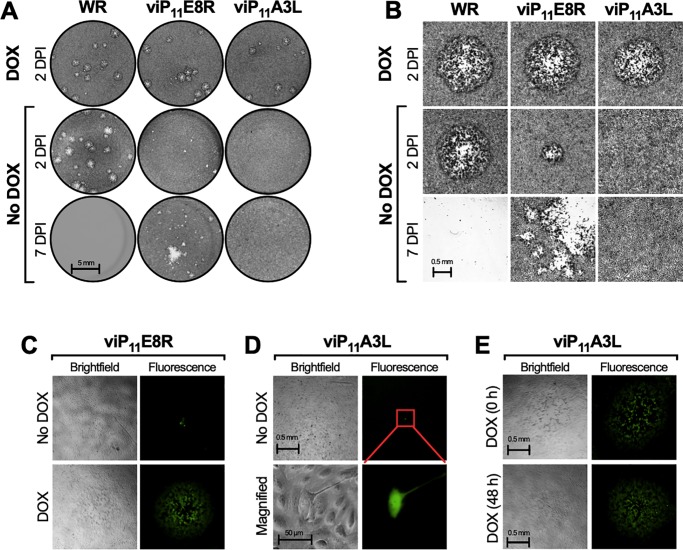
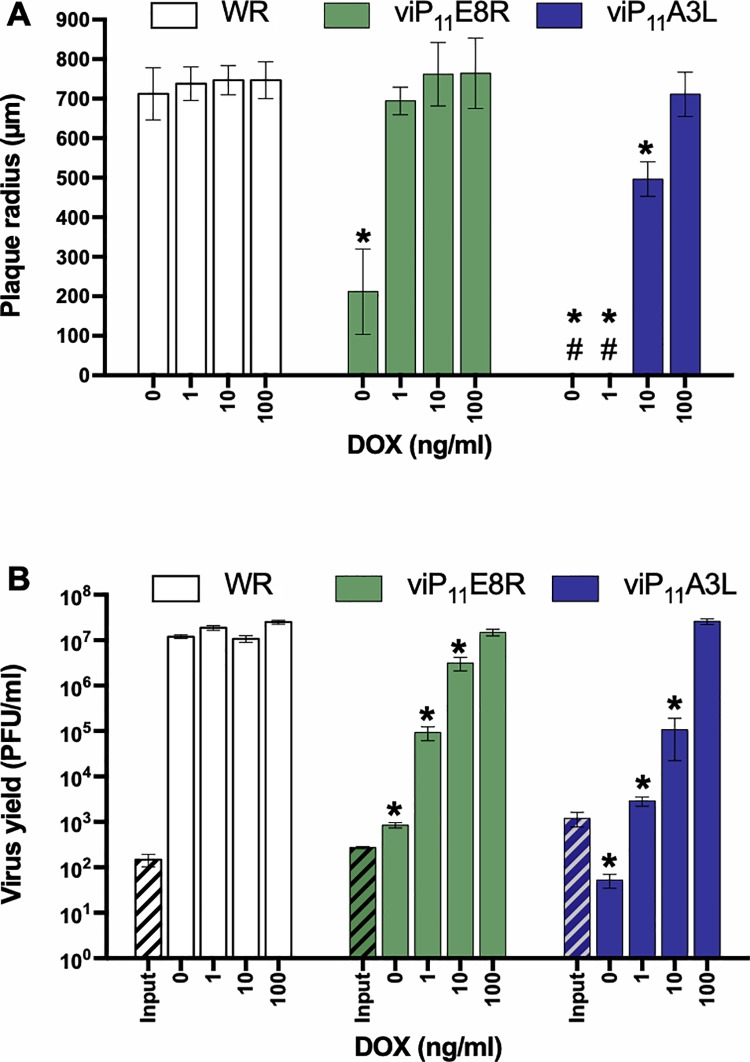
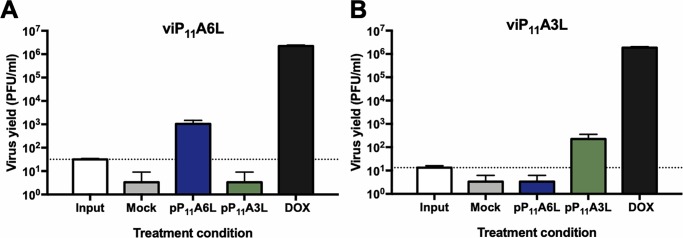
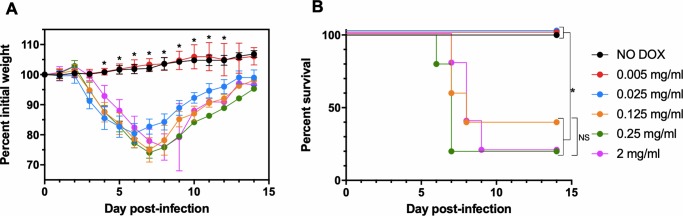
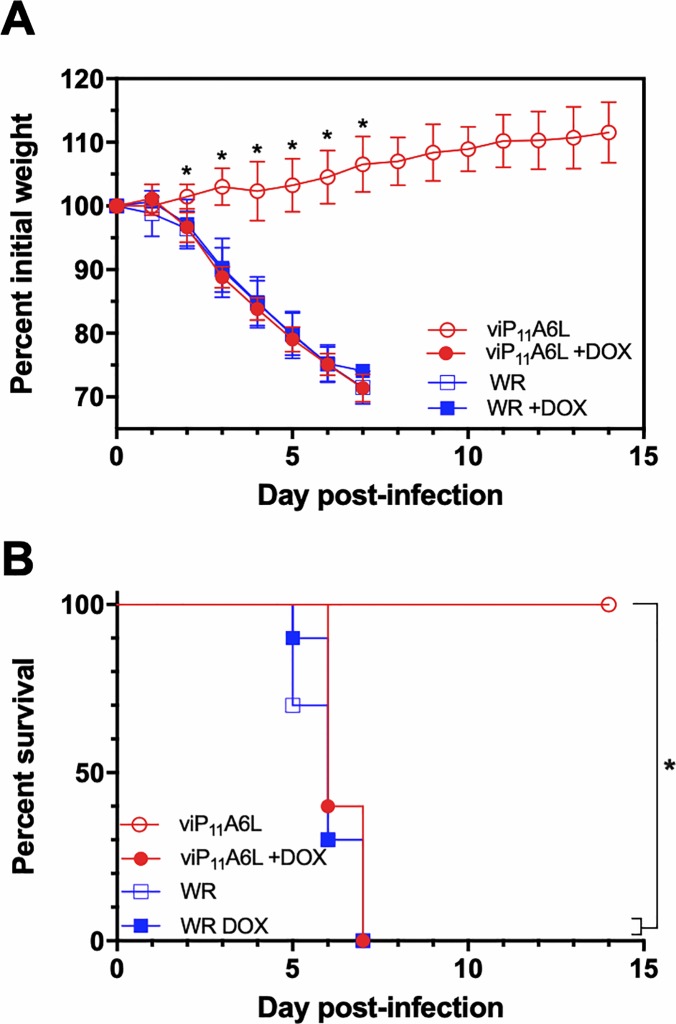
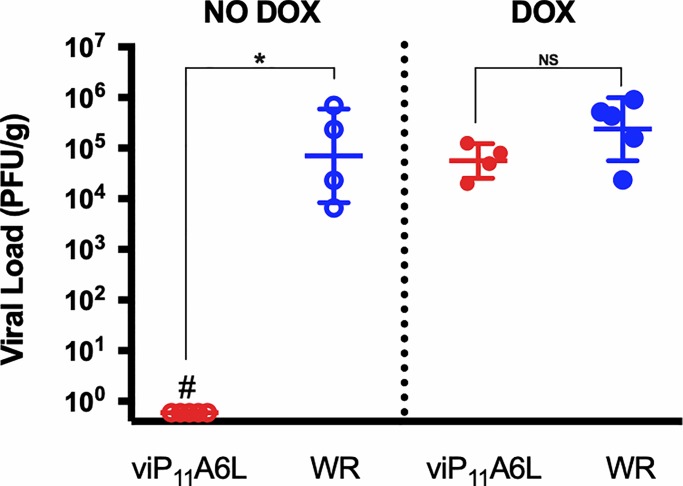
Vaccinia virus (VACV) has been used extensively as the vaccine against smallpox and as a viral vector for the development of recombinant vaccines and cancer therapies. Replication-competent, non-attenuated VACVs induce strong, long-lived humoral and cell-mediated immune responses and can be effective oncolytic vectors. However, complications from uncontrolled VACV replication in vaccinees and their close contacts can be severe, particularly in individuals with predisposing conditions. In an effort to develop replication-competent VACV vectors with improved safety, we placed VACV late genes encoding core or virion morphogenesis proteins under the control of tet operon elements to regulate their expression with tetracycline antibiotics. These replication-inducible VACVs would only express the selected genes in the presence of tetracyclines. VACVs inducibly expressing the A3L or A6L genes replicated indistinguishably from wild-type VACV in the presence of tetracyclines, whereas there was no evidence of replication in the absence of antibiotics. These outcomes were reflected in mice, where the VACV inducibly expressing the A6L gene caused weight loss and mortality equivalent to wild-type VACV in the presence of tetracyclines. In the absence of tetracyclines, mice were protected from weight loss and mortality, and viral replication was not detected. These findings indicate that replication-inducible VACVs based on the conditional expression of the A3L or A6L genes can be used for the development of safer, next-generation live VACV vectors and vaccines. The design allows for administration of replication-inducible VACV in the absence of tetracyclines (as a replication-defective vector) or in the presence of tetracyclines (as a replication-competent vector) with enhanced safety.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Prevention CfDCa. Update: multistate outbreak of monkeypox—Illinois, Indiana, Kansas, Missouri, Ohio, and Wisconsin, 2003. MMWR Morb Mortal Wkly Rep. 2003;52(27):642–6. Epub 2003/07/12. . - PubMed
-
- Fenner F. Smallpox and its eradication. Geneva: World Health Organization; 1988. xvi, 1460 p. p.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
