

A long noncoding RNA acts as a post-transcriptional regulator of heat shock protein (HSP70) synthesis in the cold hardy Diamesa tonsa under heat shock
- PMID: 32240200
- PMCID: PMC7117718
- DOI: 10.1371/journal.pone.0227172
A long noncoding RNA acts as a post-transcriptional regulator of heat shock protein (HSP70) synthesis in the cold hardy Diamesa tonsa under heat shock
Abstract
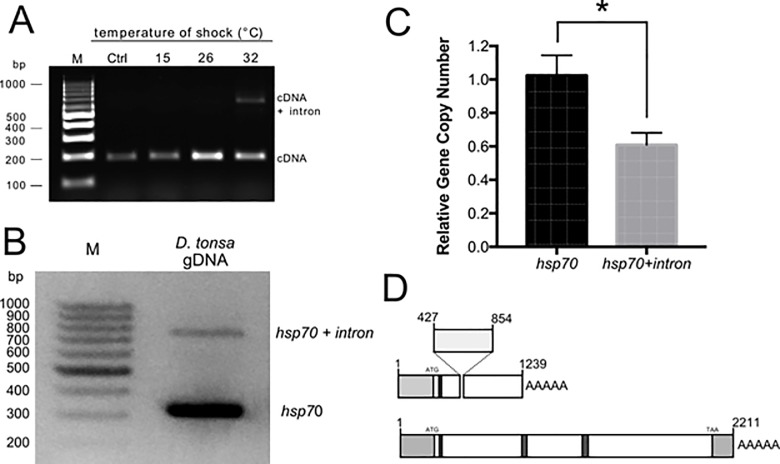
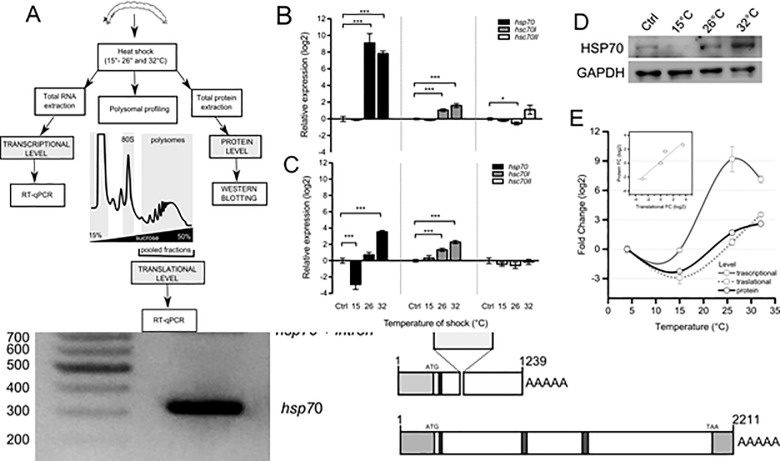
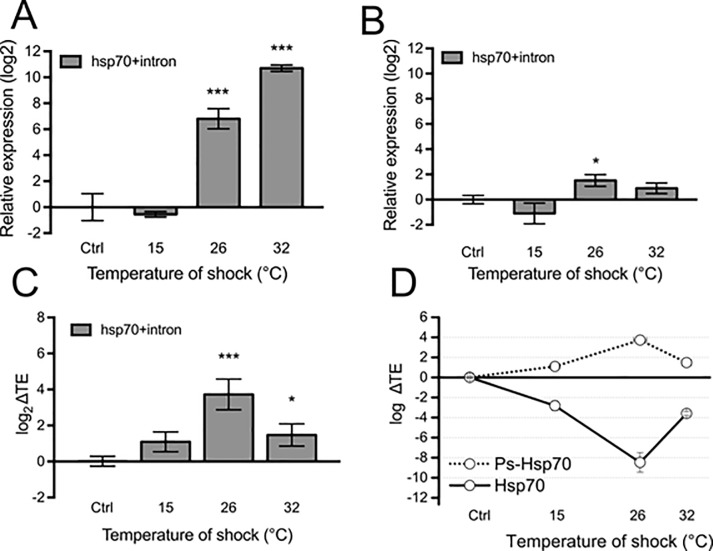
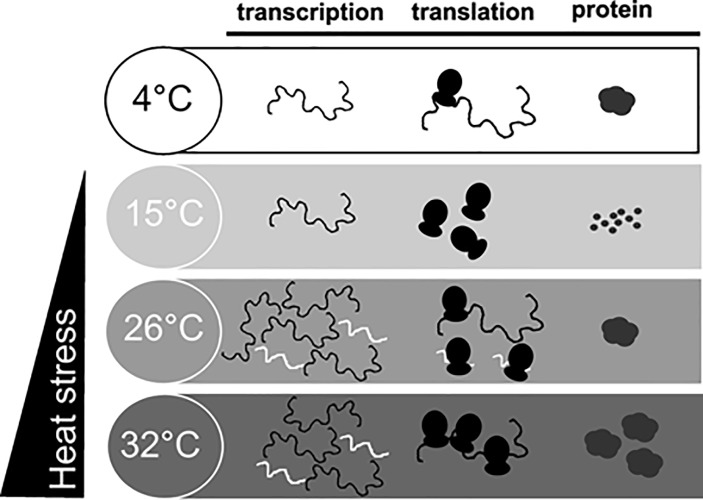
Cold stenothermal insects living in glacier-fed streams are stressed by temperature variations resulting from glacial retreat during global warming. The molecular aspects of insect response to environmental stresses remain largely unexplored. The aim of this study was to expand our knowledge of how a cold stenothermal organism controls gene expression at the transcriptional, translational, and protein level under warming conditions. Using the chironomid Diamesa tonsa as target species and a combination of RACE, qPCR, polysomal profiling, western blotting, and bioinformatics techniques, we discovered a new molecular pathway leading to previously overlooked adaptive strategies to stress. We obtained and characterized the complete cDNA sequences of three heat shock inducible 70 (hsp70) and two members of heat-shock cognate 70 (hsc70). Strikingly, we showed that a novel pseudo-hsp70 gene encoding a putative long noncoding RNA (lncRNA) which is transcribed during thermal stress, acting as a ribosome sponge to provide post-transcriptional control of HSP70 protein levels. The expression of the pseudo-hsp70 gene and its function suggest the existence of a new and unexpected mechanism to cope with thermal stress: lowering the pace of protein production to save energy and optimize resources for recovery.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Lencioni V, Spitale D. Diversity and distribution of benthic and hyporheic fauna in different stream types on an alpine glacial floodplain. Hydrobiologia. 2015;751:73–87. 10.1007/s10750-014-2172-2 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
