

Metal ion coordination delays amyloid-β peptide self-assembly by forming an aggregation-inert complex
- PMID: 32241918
- PMCID: PMC7247290
- DOI: 10.1074/jbc.RA120.012738
Metal ion coordination delays amyloid-β peptide self-assembly by forming an aggregation-inert complex
Abstract
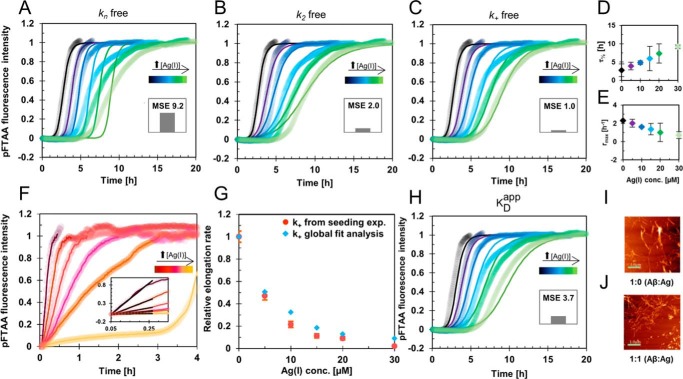
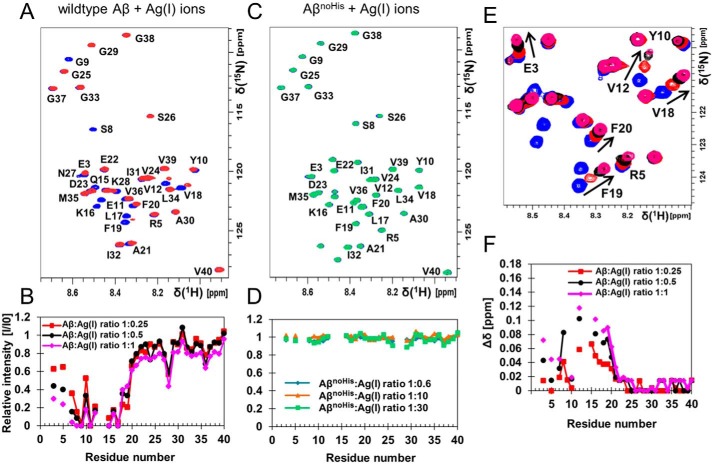
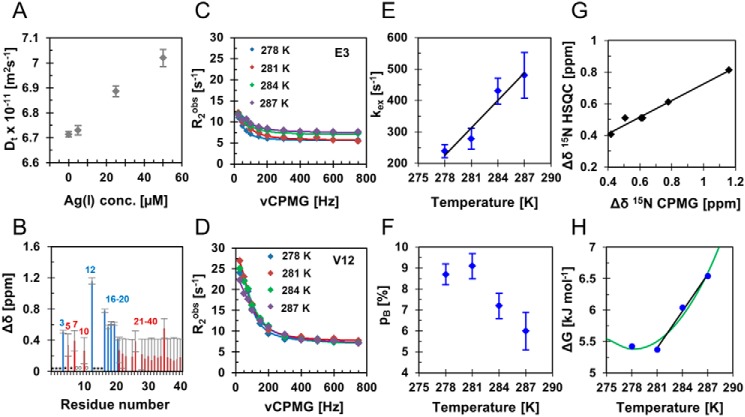
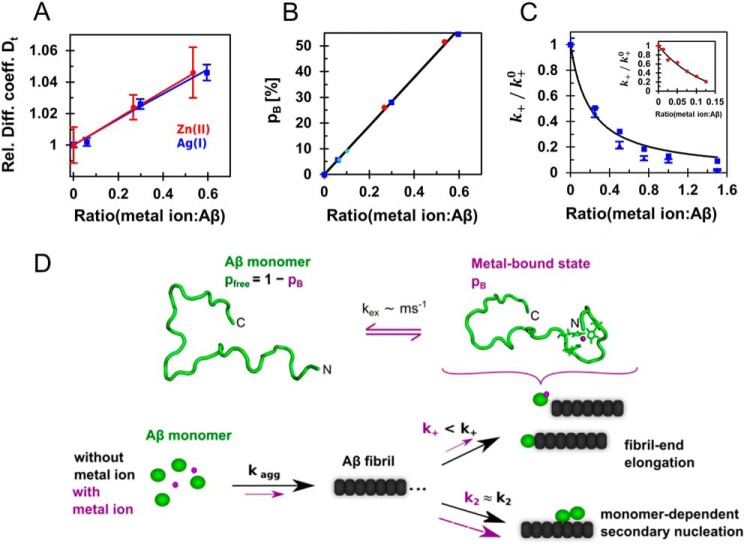
A detailed understanding of the molecular pathways for amyloid-β (Aβ) peptide aggregation from monomers into amyloid fibrils, a hallmark of Alzheimer's disease, is crucial for the development of diagnostic and therapeutic strategies. We investigate the molecular details of peptide fibrillization in vitro by perturbing this process through addition of differently charged metal ions. Here, we used a monovalent probe, the silver ion, that, similarly to divalent metal ions, binds to monomeric Aβ peptide and efficiently modulates Aβ fibrillization. On the basis of our findings, combined with our previous results on divalent zinc ions, we propose a model that links the microscopic metal-ion binding to Aβ monomers to its macroscopic impact on the peptide self-assembly observed in bulk experiments. We found that substoichiometric concentrations of the investigated metal ions bind specifically to the N-terminal region of Aβ, forming a dynamic, partially compact complex. The metal-ion bound state appears to be incapable of aggregation, effectively reducing the available monomeric Aβ pool for incorporation into fibrils. This is especially reflected in a decreased fibril-end elongation rate. However, because the bound state is significantly less stable than the amyloid state, Aβ peptides are only transiently redirected from fibril formation, and eventually almost all Aβ monomers are integrated into fibrils. Taken together, these findings unravel the mechanistic consequences of delaying Aβ aggregation via weak metal-ion binding, quantitatively linking the contributions of specific interactions of metal ions with monomeric Aβ to their effects on bulk aggregation.
Keywords: Alzheimer disease; amyloid; amyloid-beta (AB); metal; metal ion-protein interaction; monovalent ion; neurodegeneration; nuclear magnetic resonance (NMR); protein aggregation; silver; zinc.
© 2020 Wallin et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
