

De novo design of protein logic gates
- PMID: 32241946
- PMCID: PMC7397813
- DOI: 10.1126/science.aay2790
De novo design of protein logic gates
Abstract
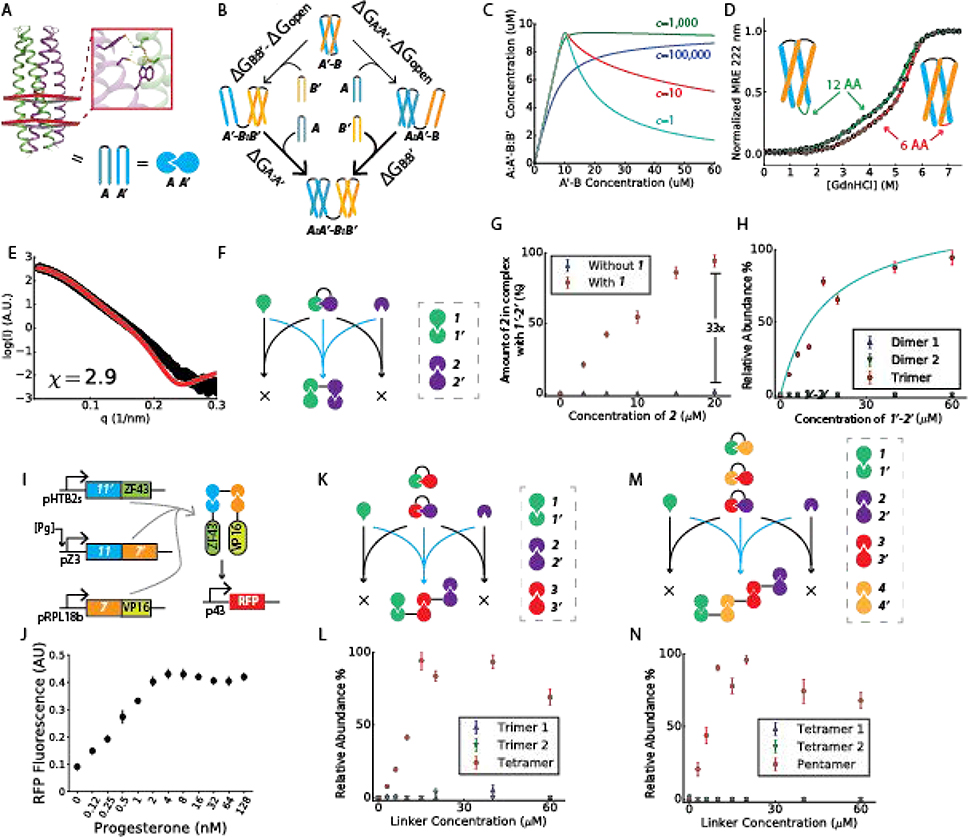
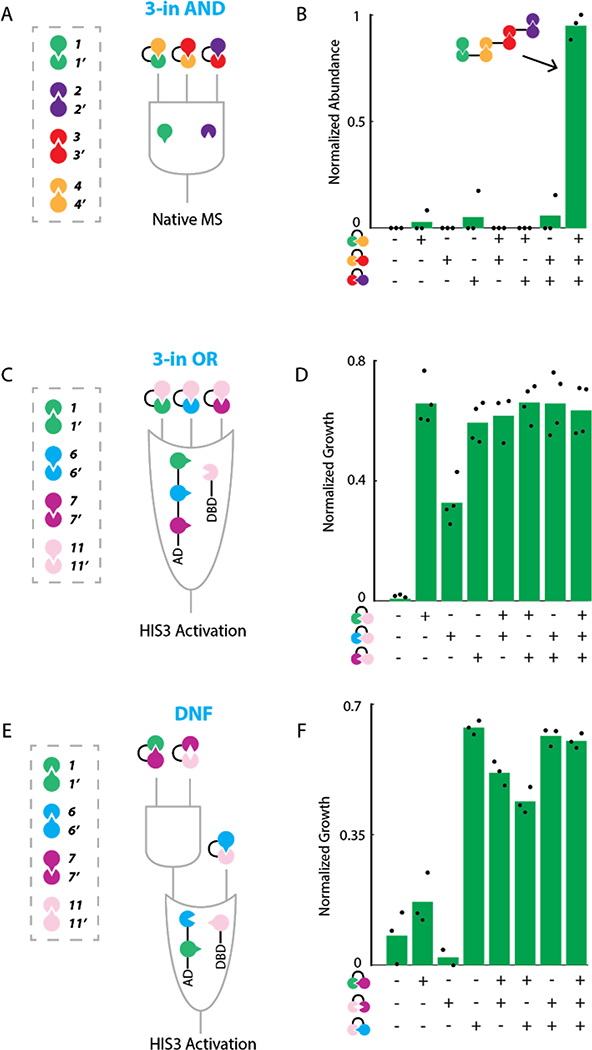
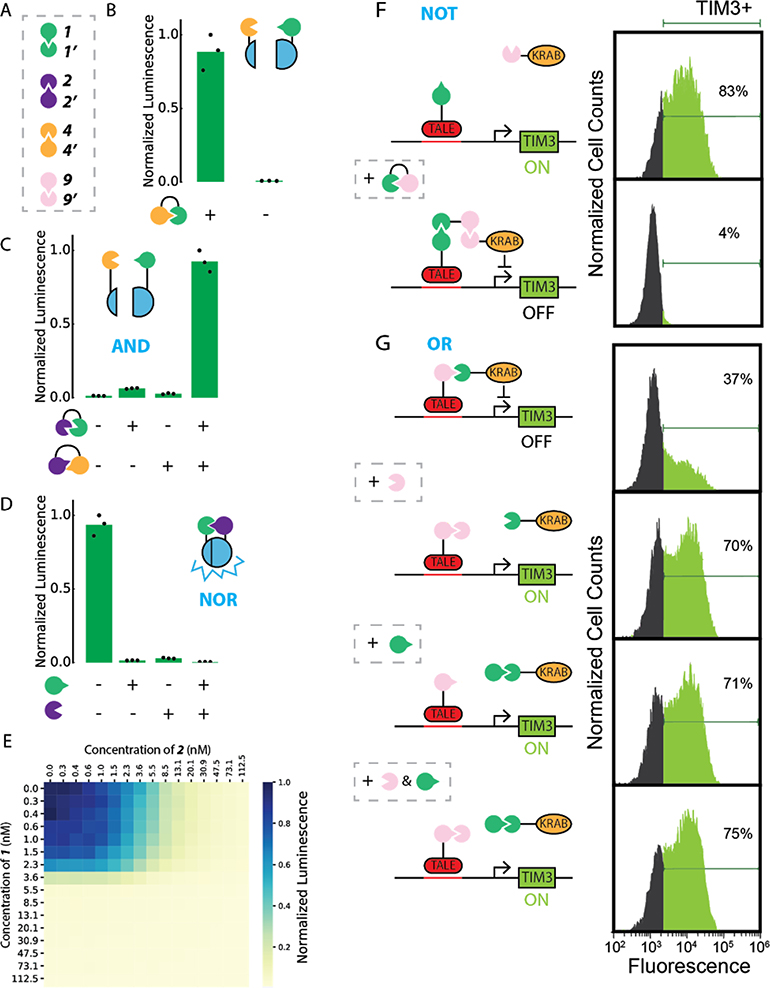
The design of modular protein logic for regulating protein function at the posttranscriptional level is a challenge for synthetic biology. Here, we describe the design of two-input AND, OR, NAND, NOR, XNOR, and NOT gates built from de novo-designed proteins. These gates regulate the association of arbitrary protein units ranging from split enzymes to transcriptional machinery in vitro, in yeast and in primary human T cells, where they control the expression of the TIM3 gene related to T cell exhaustion. Designed binding interaction cooperativity, confirmed by native mass spectrometry, makes the gates largely insensitive to stoichiometric imbalances in the inputs, and the modularity of the approach enables ready extension to three-input OR, AND, and disjunctive normal form gates. The modularity and cooperativity of the control elements, coupled with the ability to de novo design an essentially unlimited number of protein components, should enable the design of sophisticated posttranslational control logic over a wide range of biological functions.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures

Comment in
-
Designing protein logic gates.Nat Methods. 2020 Jun;17(6):565. doi: 10.1038/s41592-020-0865-1. Nat Methods. 2020. PMID: 32499622 No abstract available.
Similar articles
-
Rational design of a fusion protein to exhibit disulfide-mediated logic gate behavior.ACS Synth Biol. 2015 Apr 17;4(4):400-6. doi: 10.1021/sb500254g. Epub 2014 Aug 27. ACS Synth Biol. 2015. PMID: 25144732 Free PMC article.
-
Mammalian synthetic biology: engineering of sophisticated gene networks.J Biotechnol. 2007 Jul 15;130(4):329-45. doi: 10.1016/j.jbiotec.2007.05.014. Epub 2007 May 24. J Biotechnol. 2007. PMID: 17602777 Review.
-
Design of fast proteolysis-based signaling and logic circuits in mammalian cells.Nat Chem Biol. 2019 Feb;15(2):115-122. doi: 10.1038/s41589-018-0181-6. Epub 2018 Dec 10. Nat Chem Biol. 2019. PMID: 30531965 Free PMC article.
-
Complete all-optical processing polarization-based binary logic gates and optical processors.Opt Express. 2006 Oct 16;14(21):9879-95. doi: 10.1364/oe.14.009879. Opt Express. 2006. PMID: 19529381
-
Recognizing and engineering digital-like logic gates and switches in gene regulatory networks.Curr Opin Microbiol. 2016 Oct;33:74-82. doi: 10.1016/j.mib.2016.07.004. Epub 2016 Jul 19. Curr Opin Microbiol. 2016. PMID: 27450541 Review.
Cited by
-
The cyclic peptide G4CP2 enables the modulation of galactose metabolism in yeast by interfering with GAL4 transcriptional activity.Front Mol Biosci. 2023 Mar 1;10:1017757. doi: 10.3389/fmolb.2023.1017757. eCollection 2023. Front Mol Biosci. 2023. PMID: 36936986 Free PMC article.
-
NOT-Gated CD93 CAR T Cells Effectively Target AML with Minimized Endothelial Cross-Reactivity.Blood Cancer Discov. 2021 Sep 16;2(6):648-665. doi: 10.1158/2643-3230.BCD-20-0208. eCollection 2021 Nov. Blood Cancer Discov. 2021. PMID: 34778803 Free PMC article.
-
A rapid cell-free expression and screening platform for antibody discovery.Nat Commun. 2023 Jul 3;14(1):3897. doi: 10.1038/s41467-023-38965-w. Nat Commun. 2023. PMID: 37400446 Free PMC article.
-
AND-gated protease-activated nanosensors for programmable detection of anti-tumour immunity.Nat Nanotechnol. 2025 Mar;20(3):441-450. doi: 10.1038/s41565-024-01834-8. Epub 2025 Jan 3. Nat Nanotechnol. 2025. PMID: 39753733
-
Synthetic Biological Circuits within an Orthogonal Central Dogma.Trends Biotechnol. 2021 Jan;39(1):59-71. doi: 10.1016/j.tibtech.2020.05.013. Epub 2020 Jun 22. Trends Biotechnol. 2021. PMID: 32586633 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
