

A salvage pathway maintains highly functional respiratory complex I
- PMID: 32242014
- PMCID: PMC7118099
- DOI: 10.1038/s41467-020-15467-7
A salvage pathway maintains highly functional respiratory complex I
Abstract
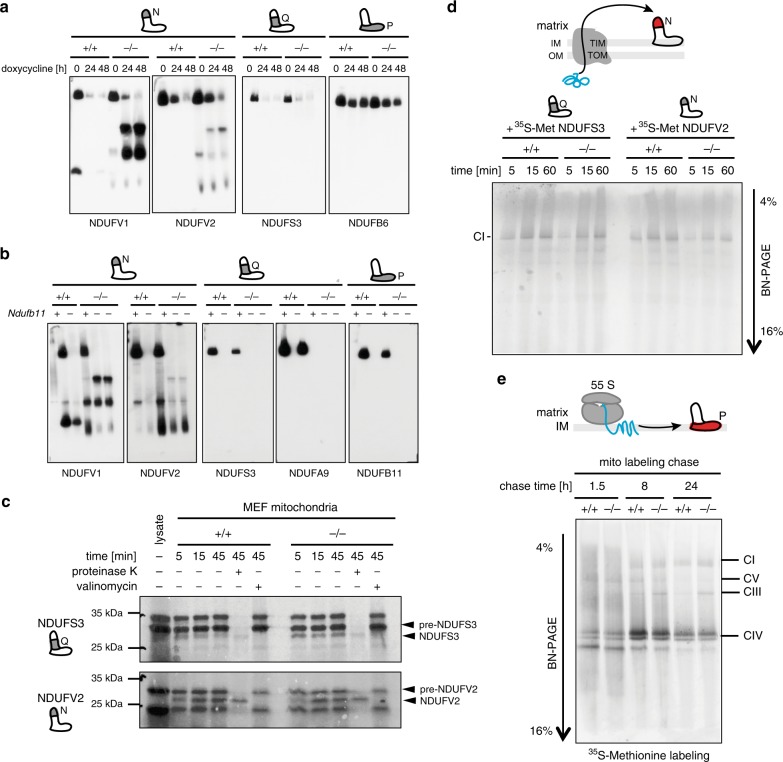
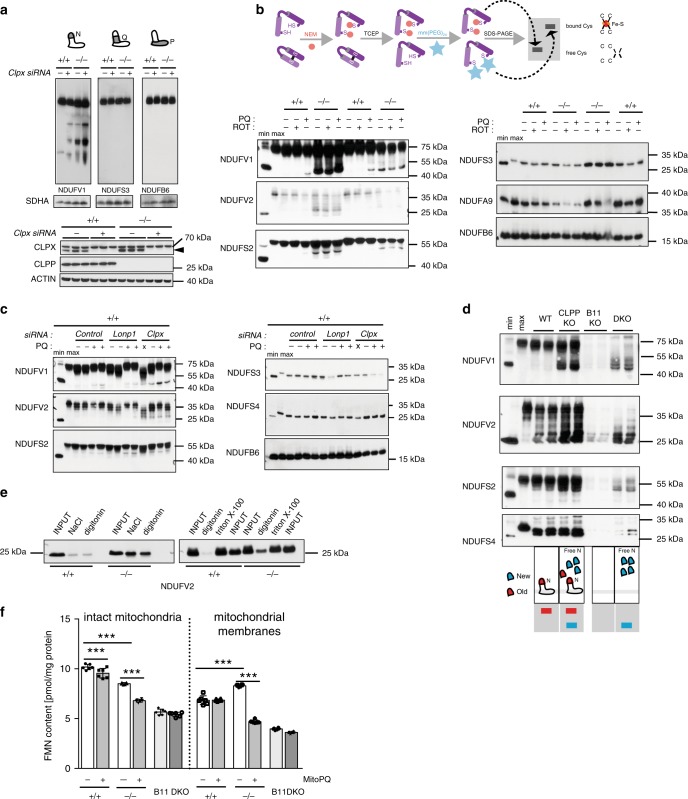
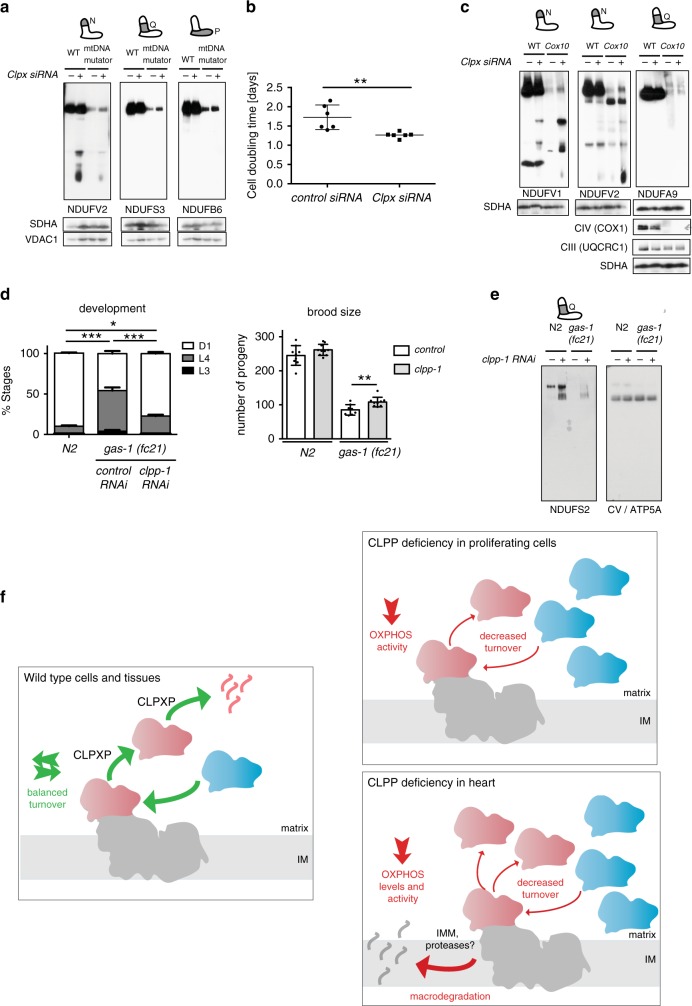
Regulation of the turnover of complex I (CI), the largest mitochondrial respiratory chain complex, remains enigmatic despite huge advancement in understanding its structure and the assembly. Here, we report that the NADH-oxidizing N-module of CI is turned over at a higher rate and largely independently of the rest of the complex by mitochondrial matrix protease ClpXP, which selectively removes and degrades damaged subunits. The observed mechanism seems to be a safeguard against the accumulation of dysfunctional CI arising from the inactivation of the N-module subunits due to attrition caused by its constant activity under physiological conditions. This CI salvage pathway maintains highly functional CI through a favorable mechanism that demands much lower energetic cost than de novo synthesis and reassembly of the entire CI. Our results also identify ClpXP activity as an unforeseen target for therapeutic interventions in the large group of mitochondrial diseases characterized by the CI instability.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Accessory subunits are integral for assembly and function of human mitochondrial complex I.Nature. 2016 Oct 6;538(7623):123-126. doi: 10.1038/nature19754. Epub 2016 Sep 14. Nature. 2016. PMID: 27626371
-
The Mitochondrial Unfoldase-Peptidase Complex ClpXP Controls Bioenergetics Stress and Metastasis.PLoS Biol. 2016 Jul 7;14(7):e1002507. doi: 10.1371/journal.pbio.1002507. eCollection 2016 Jul. PLoS Biol. 2016. PMID: 27389535 Free PMC article.
-
Five entry points of the mitochondrially encoded subunits in mammalian complex I assembly.Mol Cell Biol. 2010 Jun;30(12):3038-47. doi: 10.1128/MCB.00025-10. Epub 2010 Apr 12. Mol Cell Biol. 2010. PMID: 20385768 Free PMC article.
-
Building a complex complex: Assembly of mitochondrial respiratory chain complex I.Semin Cell Dev Biol. 2018 Apr;76:154-162. doi: 10.1016/j.semcdb.2017.08.011. Epub 2017 Aug 7. Semin Cell Dev Biol. 2018. PMID: 28797839 Review.
-
Understanding mitochondrial complex I assembly in health and disease.Biochim Biophys Acta. 2012 Jun;1817(6):851-62. doi: 10.1016/j.bbabio.2011.08.010. Epub 2011 Sep 2. Biochim Biophys Acta. 2012. PMID: 21924235 Review.
Cited by
-
The membrane domain of respiratory complex I accumulates during muscle aging in Drosophila melanogaster.Sci Rep. 2022 Dec 27;12(1):22433. doi: 10.1038/s41598-022-26414-5. Sci Rep. 2022. PMID: 36575244 Free PMC article.
-
FGF21 modulates mitochondrial stress response in cardiomyocytes only under mild mitochondrial dysfunction.Sci Adv. 2022 Apr 8;8(14):eabn7105. doi: 10.1126/sciadv.abn7105. Epub 2022 Apr 6. Sci Adv. 2022. PMID: 35385313 Free PMC article.
-
FTSH PROTEASE 3 facilitates Complex I degradation through a direct interaction with the Complex I subunit PSST.Plant Cell. 2023 Aug 2;35(8):3092-3108. doi: 10.1093/plcell/koad128. Plant Cell. 2023. PMID: 37177987 Free PMC article.
-
Impaired complex I repair causes recessive Leber's hereditary optic neuropathy.J Clin Invest. 2021 Mar 15;131(6):e138267. doi: 10.1172/JCI138267. J Clin Invest. 2021. PMID: 33465056 Free PMC article.
-
RedoxiFluor: A microplate technique to quantify target-specific protein thiol redox state in relative percentage and molar terms.Free Radic Biol Med. 2022 Mar;181:118-129. doi: 10.1016/j.freeradbiomed.2022.01.023. Epub 2022 Feb 4. Free Radic Biol Med. 2022. PMID: 35131446 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
