

Cell-intrinsic metabolic regulation of mononuclear phagocyte activation: Findings from the tip of the iceberg
- PMID: 32242952
- PMCID: PMC10911050
- DOI: 10.1111/imr.12848
Cell-intrinsic metabolic regulation of mononuclear phagocyte activation: Findings from the tip of the iceberg
Abstract
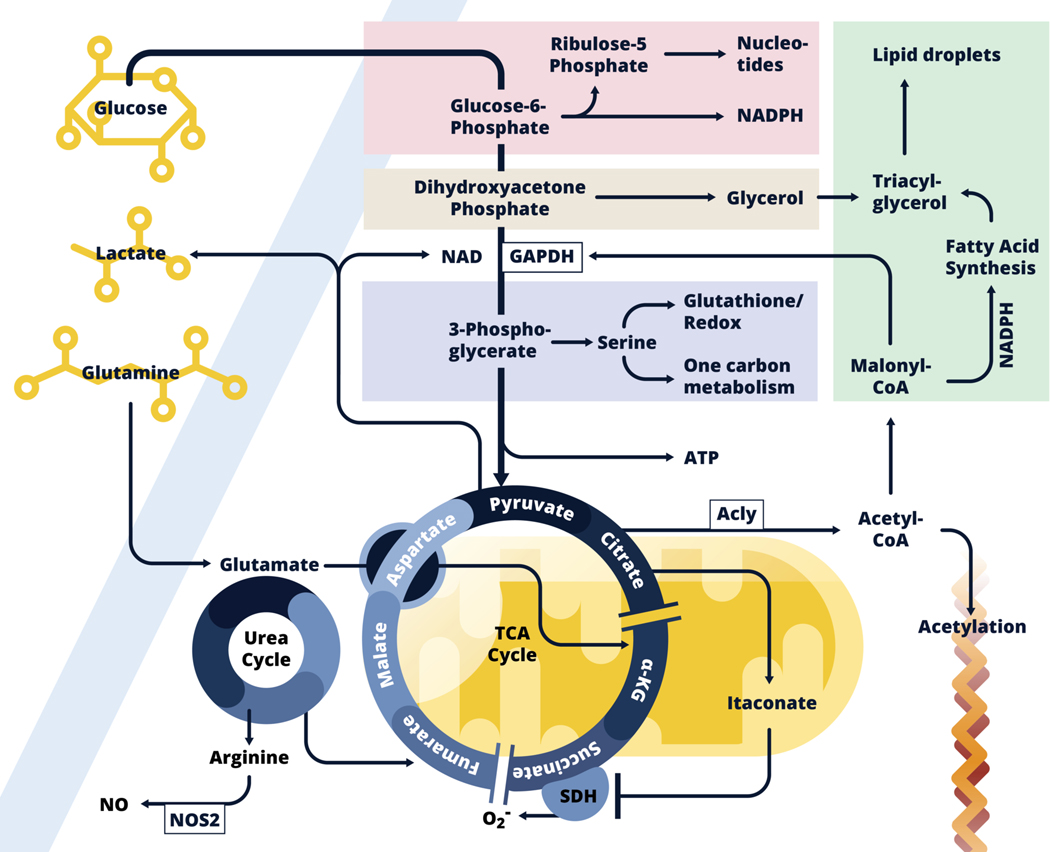
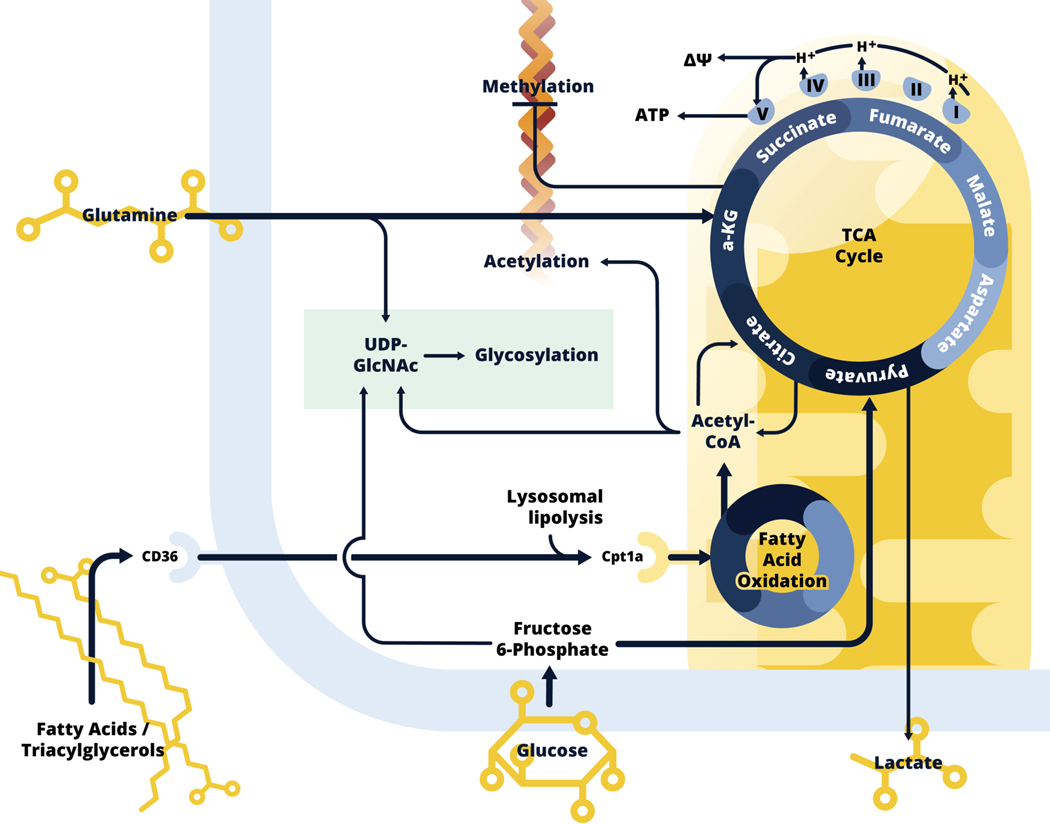
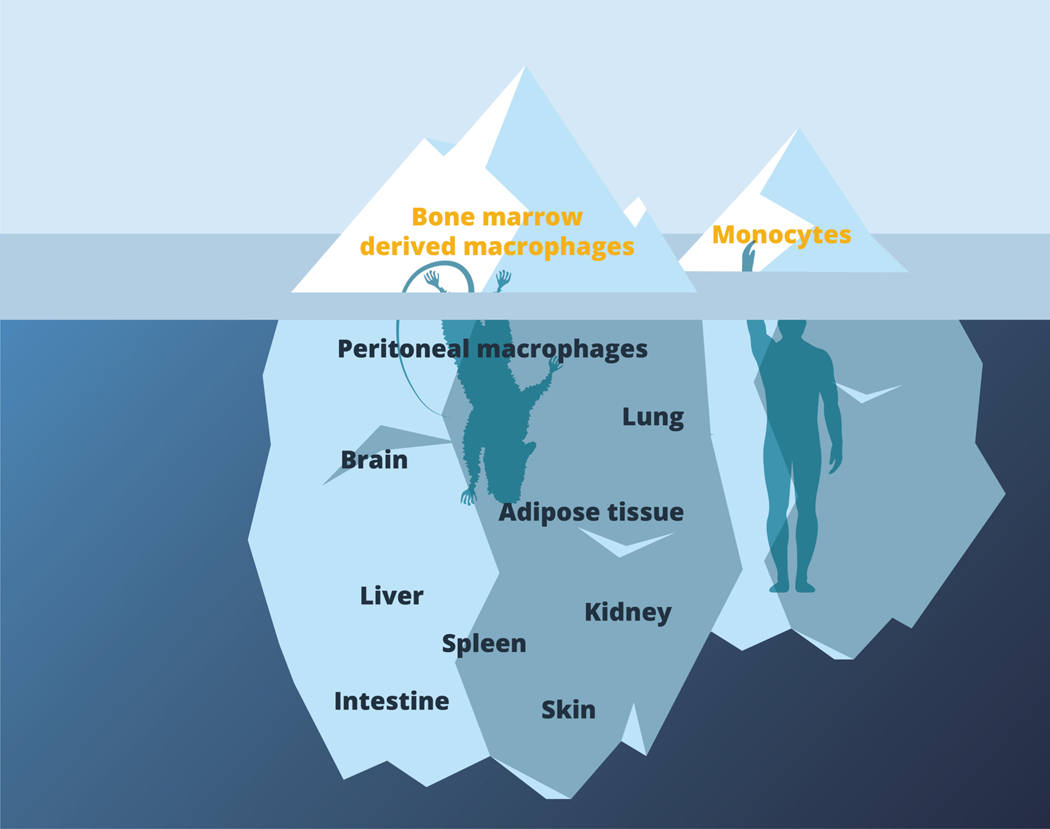
We have only recently started to appreciate the extent to which immune cell activation involves significant changes in cellular metabolism. We are now beginning to understand how commitment to specific metabolic pathways influences aspects of cellular biology that are the more usual focus of immunological studies, such as activation-induced changes in gene transcription, post-transcriptional regulation of transcription, post-translational modifications of proteins, cytokine secretion, etc. Here, we focus on metabolic reprogramming in mononuclear phagocytes downstream of stimulation with inflammatory signals (such as LPS and IFNγ) vs alternative activation signals (IL-4), with an emphasis on work on dendritic cells and macrophages from our laboratory, and related studies from others. We cover aspects of glycolysis and its branching pathways (glycogen synthesis, pentose phosphate, serine synthesis, hexose synthesis, and glycerol 3 phosphate shuttle), the tricarboxylic acid pathway, fatty acid synthesis and oxidation, and mitochondrial biology. Although our understanding of the metabolism of mononuclear phagocytes has progressed significantly over the last 10 years, major challenges remain, including understanding the effects of tissue residence on metabolic programming related to cellular activation, and the translatability of findings from mouse to human biology.
Keywords: IL-4; LPS; TCA cycle; alternative activation; cytokines; dendritic cells; glycolysis; immunometabolism; inflammation; macrophages; metabolism; mitochondria.
© 2020 The Authors. Immunological Reviews published by John Wiley & Sons Ltd.
Conflict of interest statement
Conflict of interest
EJP is a founder of Rheos Medicines.
Figures
References
-
- Kapsenberg ML. Dendritic-cell control of pathogen-driven T-cell polarization. Nat Rev Immunol. 2003;3(12):984–93. - PubMed
-
- Joffre O, Nolte MA, Spörri R, Sousa CRE. Inflammatory signals in dendritic cell activation and the induction of adaptive immunity. Immunol Rev. 2009;227(1):234–47. - PubMed
-
- Jones RG, Thompson CB. Revving the engine: signal transduction fuels T cell activation. Immunity. 2007. Aug;27(2):173–8. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
