

Metabolic Reprogramming Promotes Neural Crest Migration via Yap/Tead Signaling
- PMID: 32243782
- PMCID: PMC7236757
- DOI: 10.1016/j.devcel.2020.03.005
Metabolic Reprogramming Promotes Neural Crest Migration via Yap/Tead Signaling
Abstract
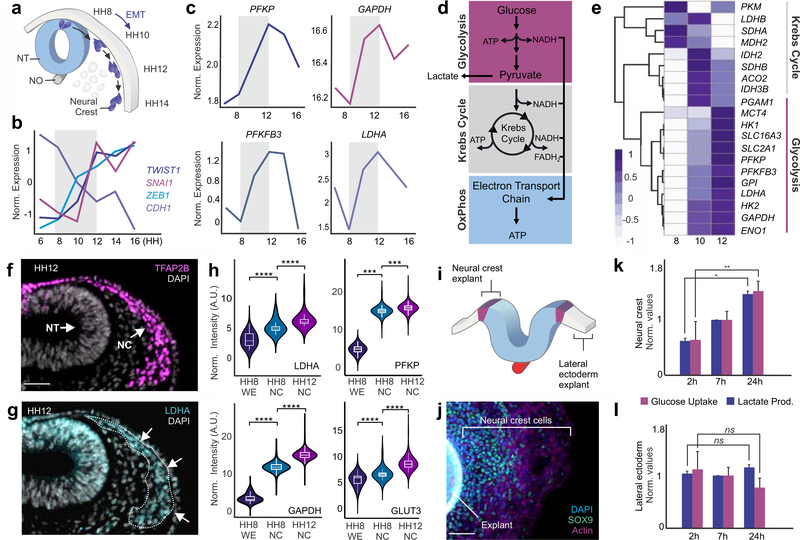
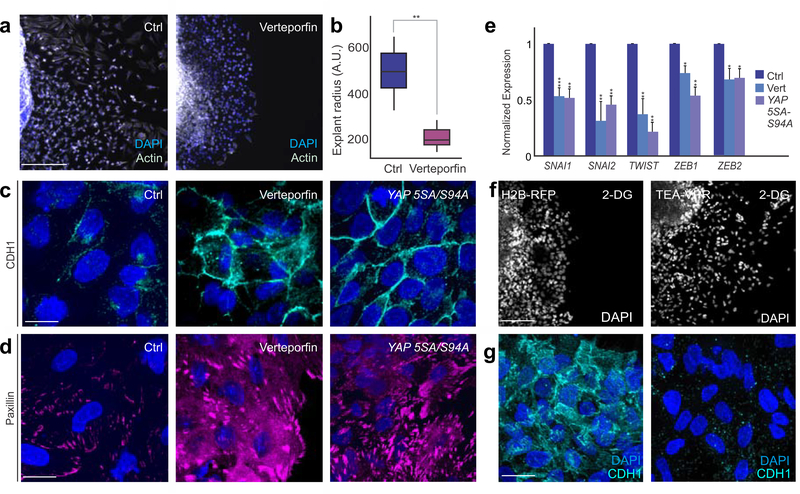
The Warburg effect is one of the metabolic hallmarks of cancer cells, characterized by enhanced glycolysis even under aerobic conditions. This physiological adaptation is associated with metastasis , but we still have a superficial understanding of how it affects cellular processes during embryonic development. Here we report that the neural crest, a migratory stem cell population in vertebrate embryos, undergoes an extensive metabolic remodeling to engage in aerobic glycolysis prior to delamination. This increase in glycolytic flux promotes Yap/Tead signaling, which activates the expression of a set of transcription factors to drive epithelial-to-mesenchymal transition. Our results demonstrate how shifts in carbon metabolism can trigger the gene regulatory circuits that control complex cell behaviors. These findings support the hypothesis that the Warburg effect is a precisely regulated developmental mechanism that is anomalously reactivated during tumorigenesis and metastasis.
Keywords: Warburg effect; Yap/Tead signaling; cell metabolism; cell migration; epithelial to mesenchymal transition; glycolysis; neural crest.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures





Comment in
-
Glucose Metabolism Takes Center Stage in Epithelial-Mesenchymal Plasticity.Dev Cell. 2020 Apr 20;53(2):133-135. doi: 10.1016/j.devcel.2020.03.021. Dev Cell. 2020. PMID: 32315608
References
-
- Barban S, and Schulze HO (1961). The effects of 2-deoxyglucose on the growth and metabolism of cultured human cells. J Biol Chem 236, 1887–1890. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
