

Liver fluke granulin promotes extracellular vesicle-mediated crosstalk and cellular microenvironment conducive to cholangiocarcinoma
- PMID: 32244128
- PMCID: PMC7118280
- DOI: 10.1016/j.neo.2020.02.004
Liver fluke granulin promotes extracellular vesicle-mediated crosstalk and cellular microenvironment conducive to cholangiocarcinoma
Abstract
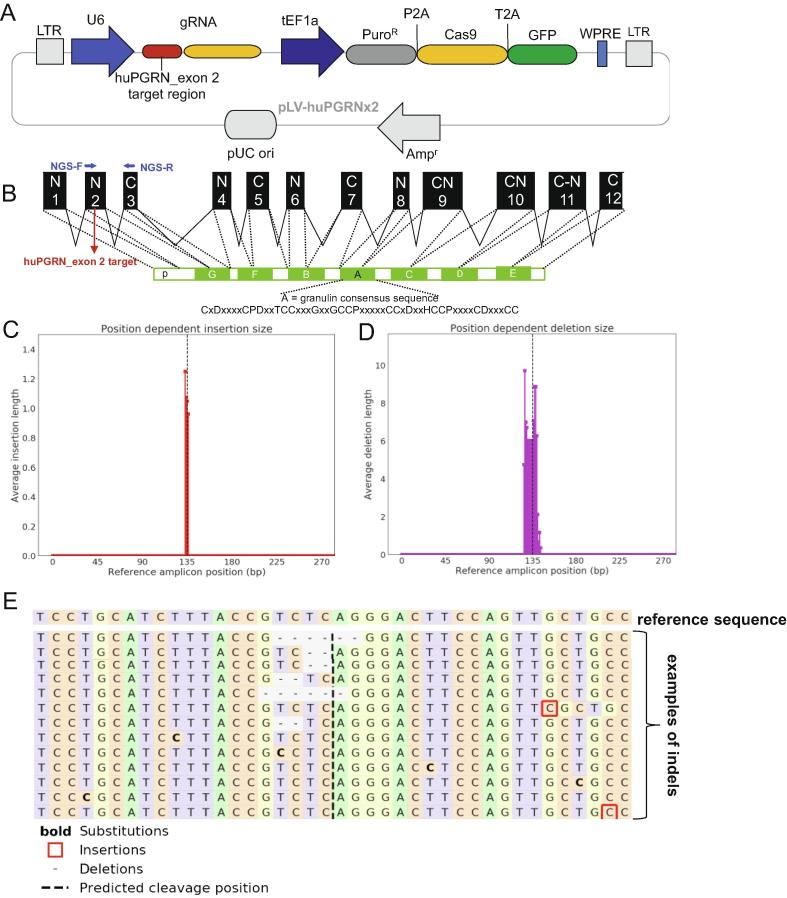
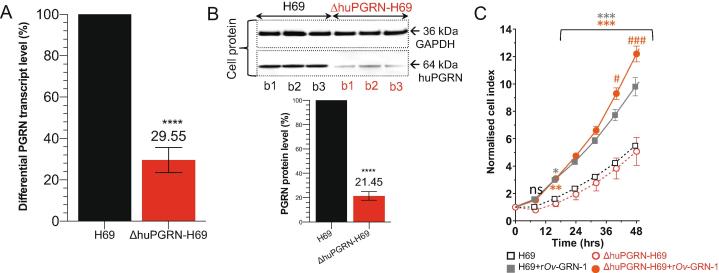
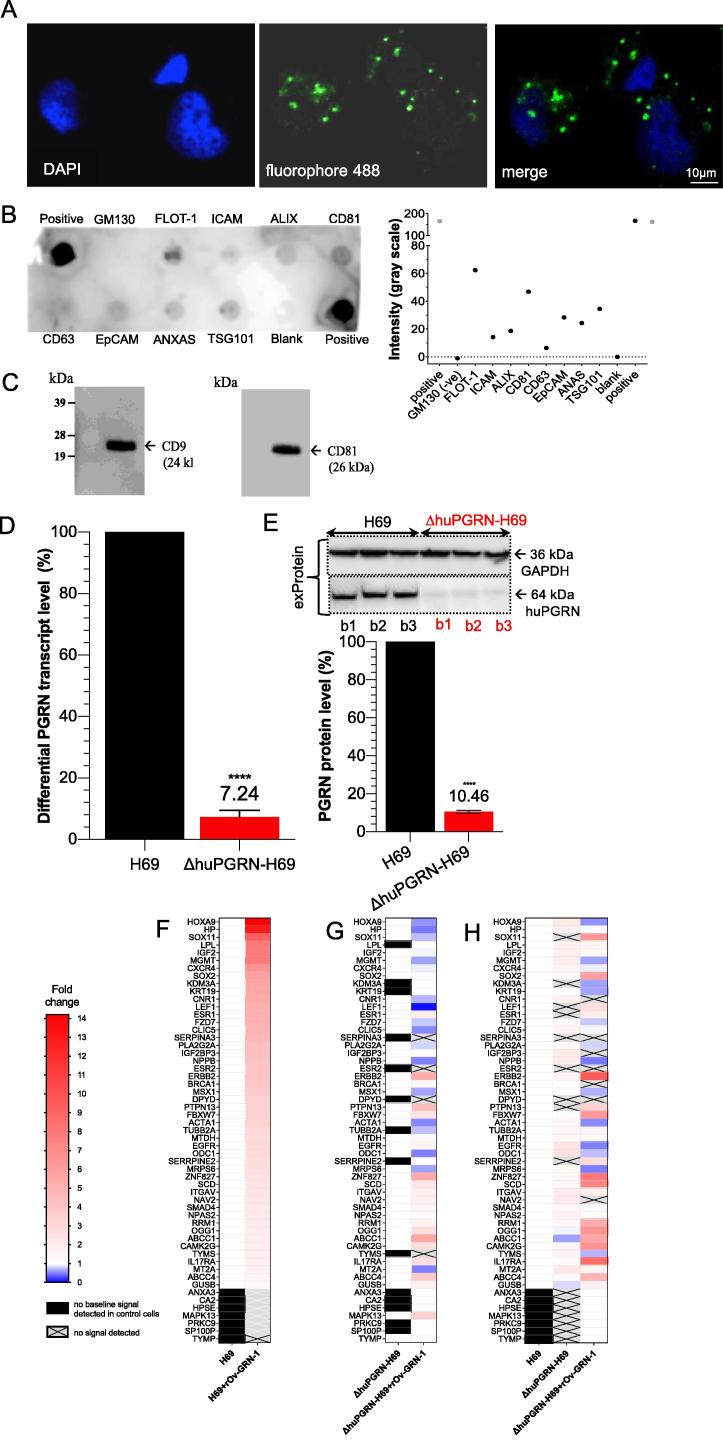
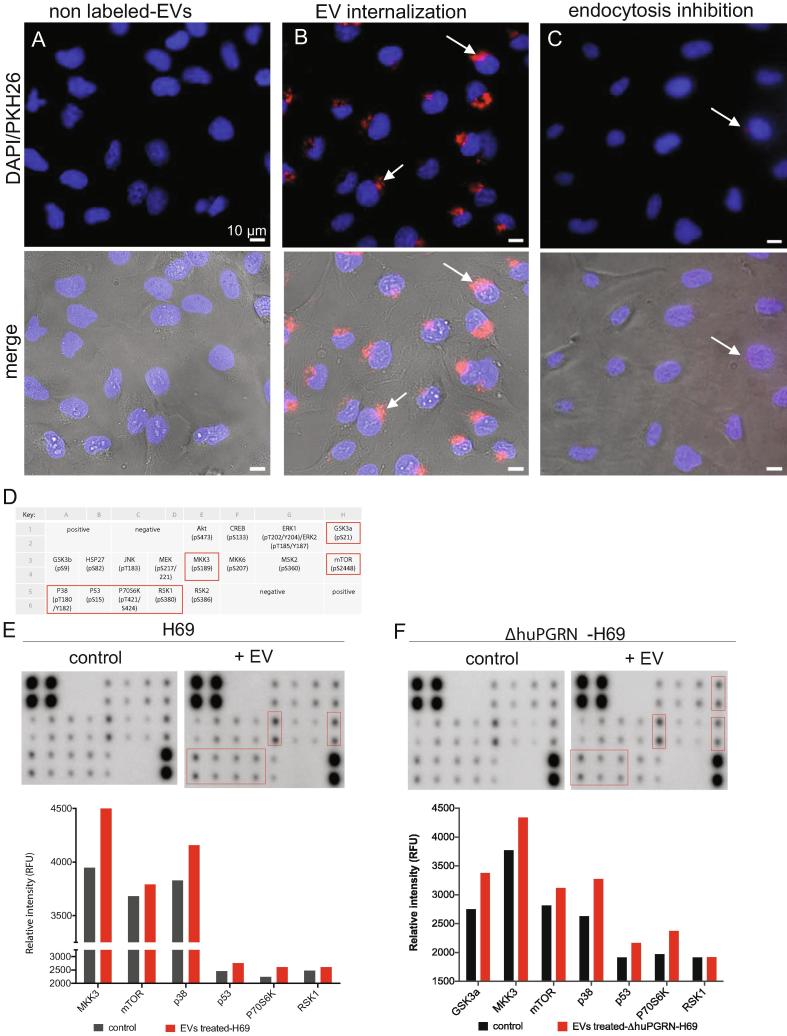
Crosstalk between malignant and neighboring cells contributes to tumor growth. In East Asia, infection with the liver fluke is a major risk factor for cholangiocarcinoma (CCA). The liver fluke Opisthorchis viverrini secretes a growth factor termed liver fluke granulin, a homologue of the human progranulin, which contributes significantly to biliary tract fibrosis and morbidity. Here, extracellular vesicle (EV)-mediated transfer of mRNAs from human cholangiocytes to naïve recipient cells was investigated following exposure to liver fluke granulin. To minimize the influence of endogenous progranulin, its cognate gene was inactivated using CRISPR/Cas9-based gene knock-out. Several progranulin-depleted cell lines, termed ΔhuPGRN-H69, were established. These lines exhibited >80% reductions in levels of specific transcript and progranulin, both in gene-edited cells and within EVs released by these cells. Profiles of extracellular vesicle RNAs (evRNA) from ΔhuPGRN-H69 for CCA-associated characteristics revealed a paucity of transcripts for estrogen- and Wnt-signaling pathways, peptidase inhibitors and tyrosine phosphatase related to cellular processes including oncogenic transformation. Several CCA-specific evRNAs including MAPK/AKT pathway members were induced by exposure to liver fluke granulin. By comparison, estrogen, Wnt/PI3K and TGF signaling and other CCA pathway mRNAs were upregulated in wild type H69 cells exposed to liver fluke granulin. Of these, CCA-associated evRNAs modified the CCA microenvironment in naïve cells co-cultured with EVs from ΔhuPGRN-H69 cells exposed to liver fluke granulin, and induced translation of MAPK phosphorylation related-protein in naïve recipient cells in comparison with control recipient cells. Exosome-mediated crosstalk in response to liver fluke granulin promoted a CCA-specific program through MAPK pathway which, in turn, established a CCA-conducive disposition.
Keywords: Cellular crosstalk; Extracellular vesicle; Liver fluke granulin; Opisthorchis viverrini.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Khan A.S., Dageforde L.A. Cholangiocarcinoma. Surg Clin North Am. 2019;99:315–335. - PubMed
-
- Fedorova O.S., Kovshirina Y.V., Kovshirina A.E., Fedotova M.M., Deev I.A., Petrovskiy F.I., Filimonov A.V., Dmitrieva A.I., Kudyakov L.A., Saltykova I.V., Odermatt P., Ogorodova L.M. Opisthorchis felineus infection and cholangiocarcinoma in the Russian Federation: a review of medical statistics. Parasitol Int. 2017;66:365–371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
