

The Multifactorial Role of PARP-1 in Tumor Microenvironment
- PMID: 32245040
- PMCID: PMC7140056
- DOI: 10.3390/cancers12030739
The Multifactorial Role of PARP-1 in Tumor Microenvironment
Abstract
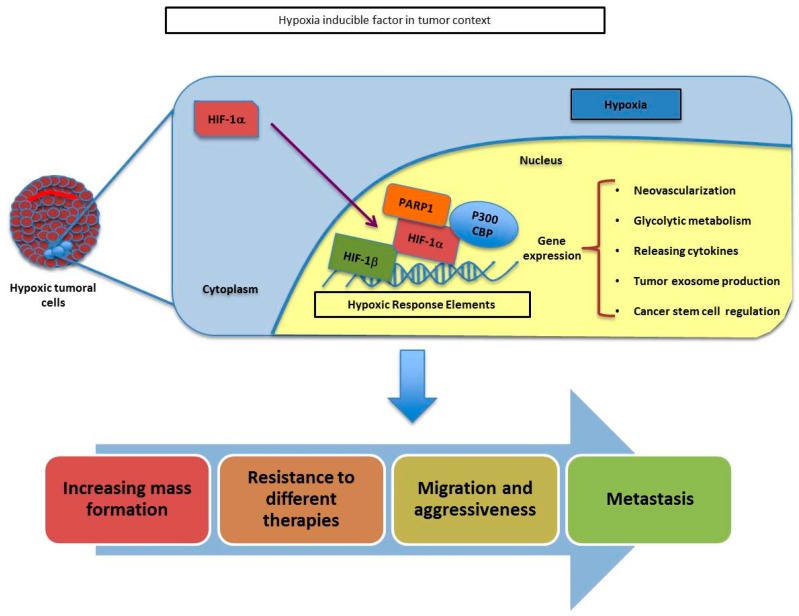
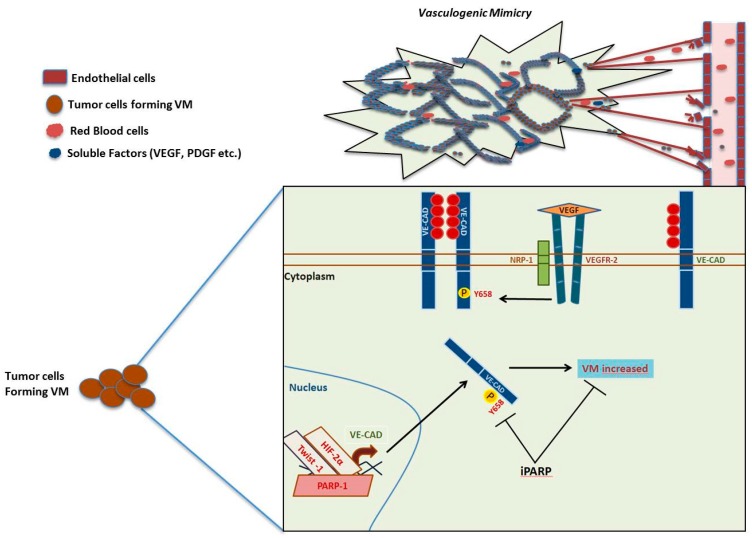
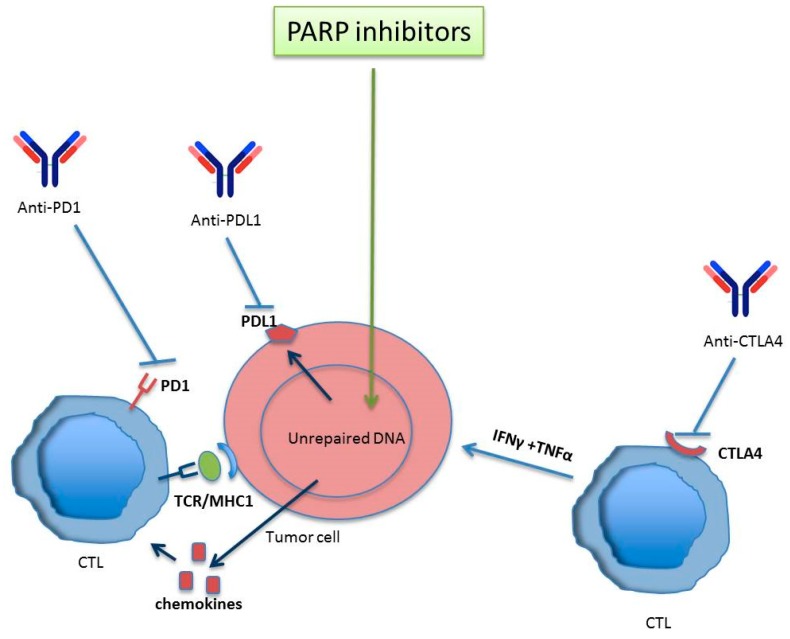
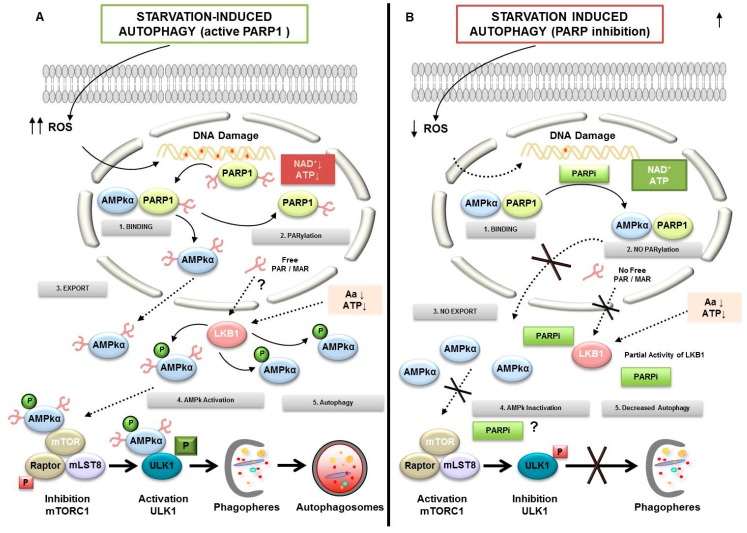
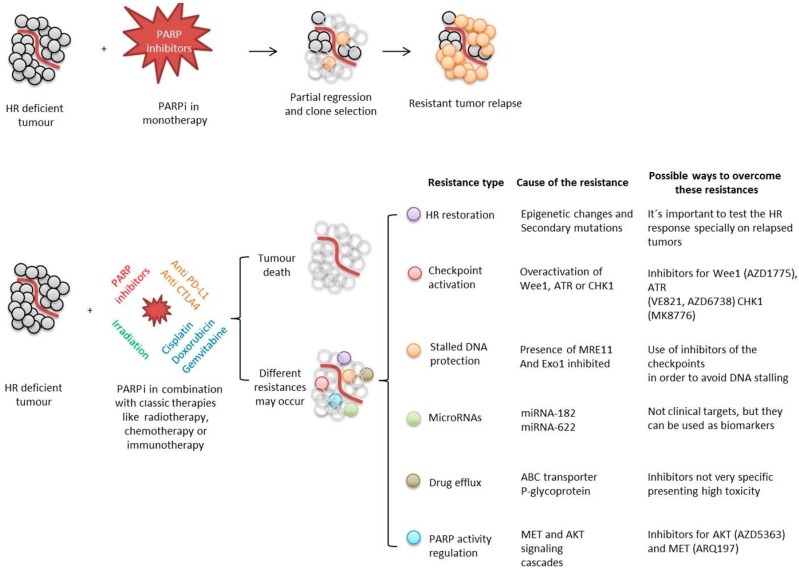
Poly(ADP-ribose) polymerases (PARPs), represent a family of 17 proteins implicated in a variety of cell functions; some of them possess the enzymatic ability to synthesize and attach poly (ADP-ribose) (also known as PAR) to different protein substrates by a post-translational modification; PARPs are key components in the cellular response to stress with consequences for different physiological and pathological events, especially during neoplasia. In recent years, using PARP inhibitors as antitumor agents has raised new challenges in understanding their role in tumor biology. Notably, the function of PARPs and PAR in the dynamic of tumor microenvironment is only starting to be understood. In this review, we summarized the conclusions arising from recent studies on the interaction between PARPs, PAR and key features of tumor microenvironment such as hypoxia, autophagy, tumor initiating cells, angiogenesis and cancer-associated immune response.
Keywords: PARP inhibitors; PARPs; PARylation; Tumor microenvironment; autophagy; hypoxia.
Conflict of interest statement
The authors declare “no conflict of interests”.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous
