

eIF4E and Interactors from Unicellular Eukaryotes
- PMID: 32245232
- PMCID: PMC7139794
- DOI: 10.3390/ijms21062170
eIF4E and Interactors from Unicellular Eukaryotes
Abstract
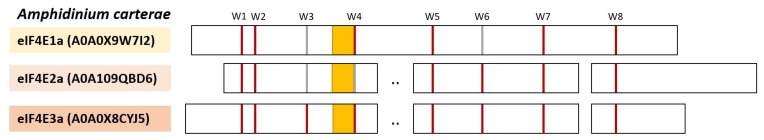
eIF4E, the mRNA cap-binding protein, is well known as a general initiation factor allowing for mRNA-ribosome interaction and cap-dependent translation in eukaryotic cells. In this review we focus on eIF4E and its interactors in unicellular organisms such as yeasts and protozoan eukaryotes. In a first part, we describe eIF4Es from yeast species such as Saccharomyces cerevisiae, Candida albicans, and Schizosaccharomyces pombe. In the second part, we will address eIF4E and interactors from parasite unicellular species-trypanosomatids and marine microorganisms-dinoflagellates. We propose that different strategies have evolved during evolution to accommodate cap-dependent translation to differing requirements. These evolutive "adjustments" involve various forms of eIF4E that are not encountered in all microorganismic species. In yeasts, eIF4E interactors, particularly p20 and Eap1 are found exclusively in Saccharomycotina species such as S. cerevisiae and C. albicans. For protozoan parasites of the Trypanosomatidae family beside a unique cap4-structure located at the 5'UTR of all mRNAs, different eIF4Es and eIF4Gs are active depending on the life cycle stage of the parasite. Additionally, an eIF4E-interacting protein has been identified in Leishmania major which is important for switching from promastigote to amastigote stages. For dinoflagellates, little is known about the structure and function of the multiple and diverse eIF4Es that have been identified thanks to widespread sequencing in recent years.
Keywords: eIF4E; eIF4E-interactors; protozoan parasites; translation; yeast.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
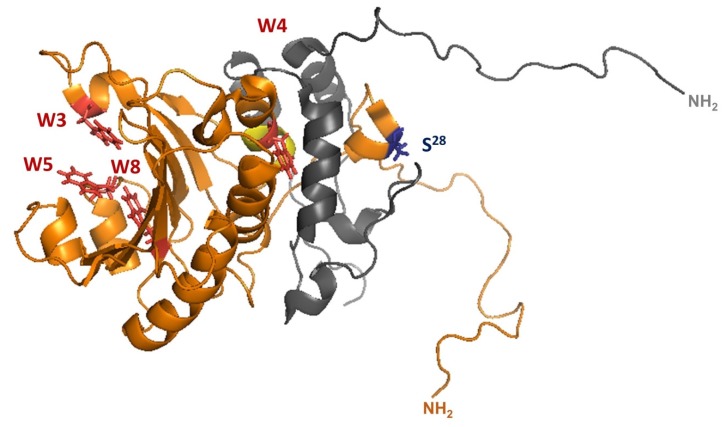
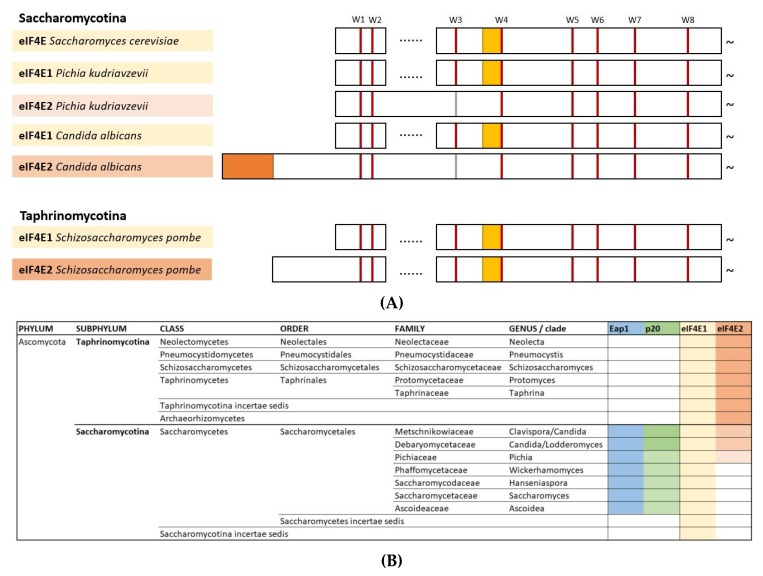
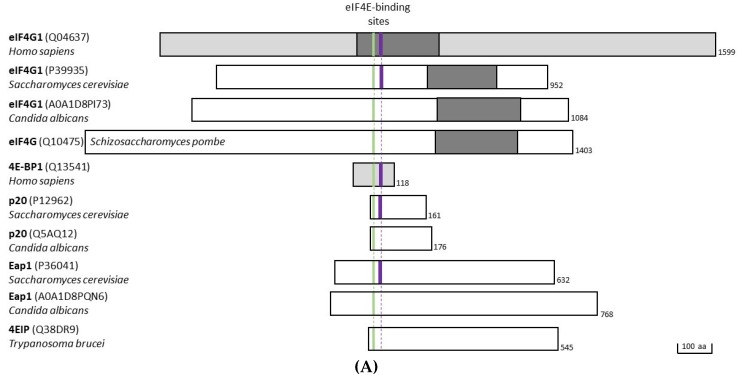

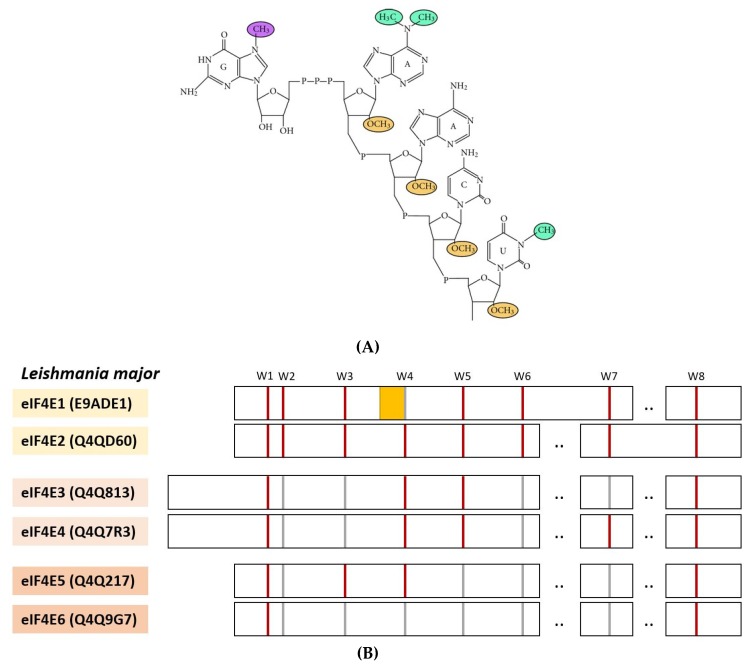
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
