

Regulatory T cells confer a circadian signature on inflammatory arthritis
- PMID: 32245954
- PMCID: PMC7125185
- DOI: 10.1038/s41467-020-15525-0
Regulatory T cells confer a circadian signature on inflammatory arthritis
Abstract
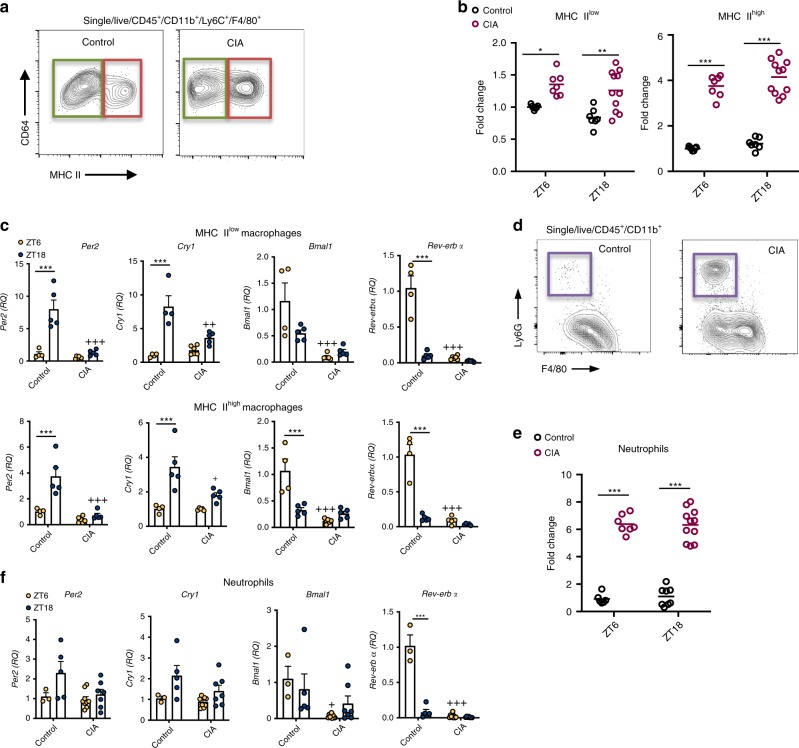
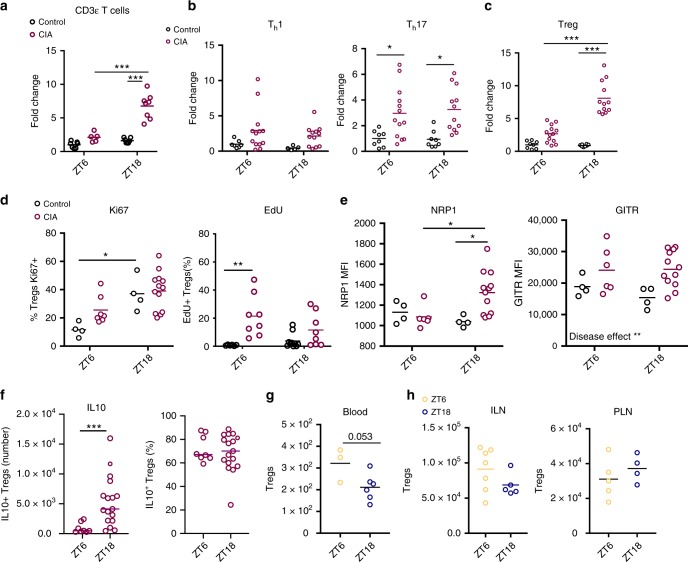
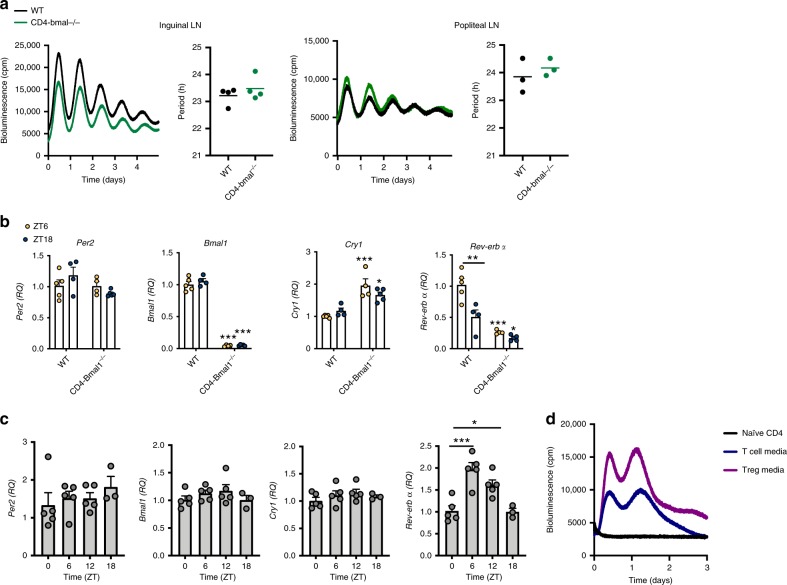
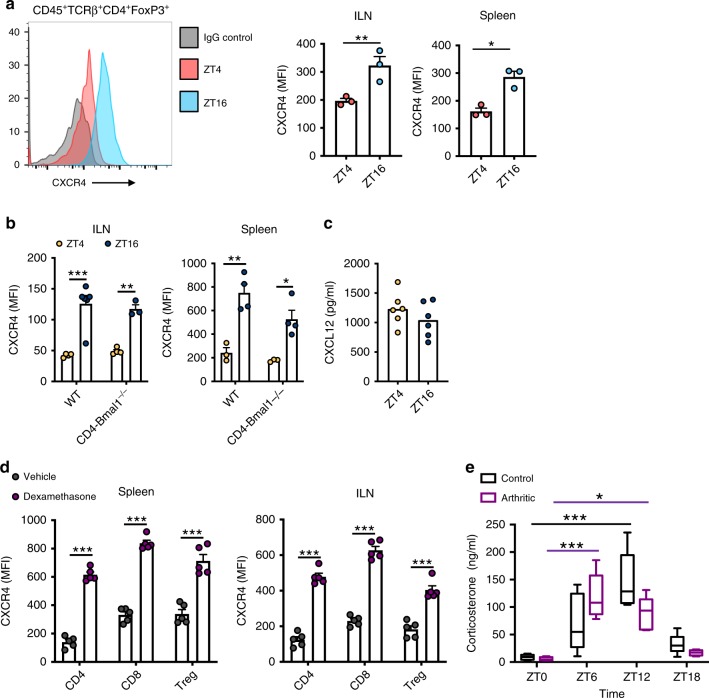
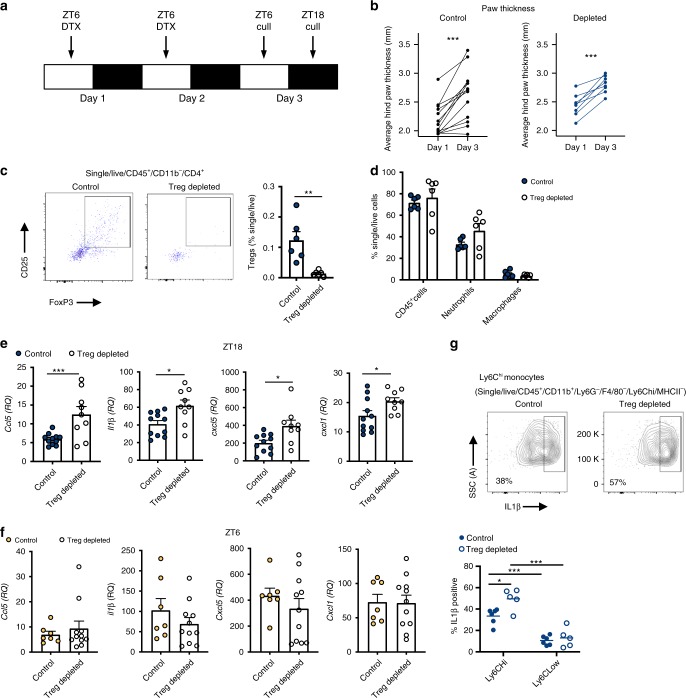
The circadian clock is an intrinsic oscillator that imparts 24 h rhythms on immunity. This clock drives rhythmic repression of inflammatory arthritis during the night in mice, but mechanisms underlying this effect are not clear. Here we show that the amplitude of intrinsic oscillators within macrophages and neutrophils is limited by the chronic inflammatory environment, suggesting that rhythms in inflammatory mediators might not be a direct consequence of intrinsic clocks. Anti-inflammatory regulatory T (Treg) cells within the joints show diurnal variation, with numbers peaking during the nadir of inflammation. Furthermore, the anti-inflammatory action of Treg cells on innate immune cells contributes to the night-time repression of inflammation. Treg cells do not seem to have intrinsic circadian oscillators, suggesting that rhythmic function might be a consequence of external signals. These data support a model in which non-rhythmic Treg cells are driven to rhythmic activity by systemic signals to confer a circadian signature to chronic arthritis.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
