

CD300lf is the primary physiologic receptor of murine norovirus but not human norovirus
- PMID: 32251490
- PMCID: PMC7162533
- DOI: 10.1371/journal.ppat.1008242
CD300lf is the primary physiologic receptor of murine norovirus but not human norovirus
Abstract
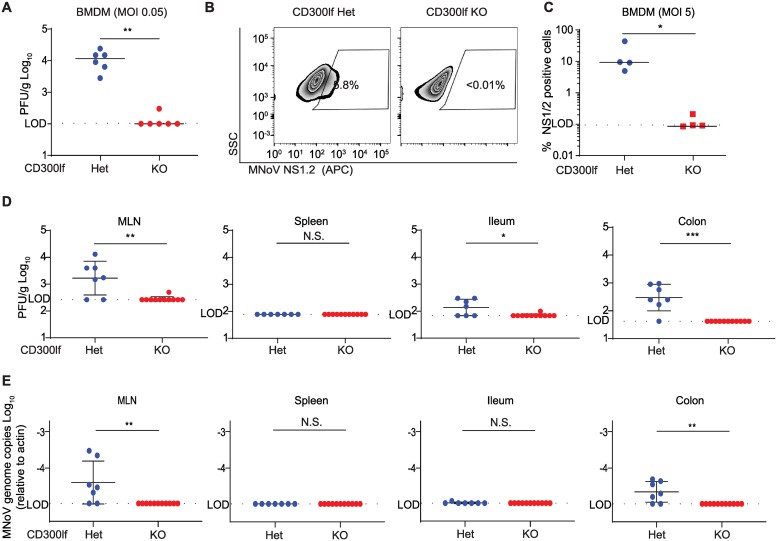
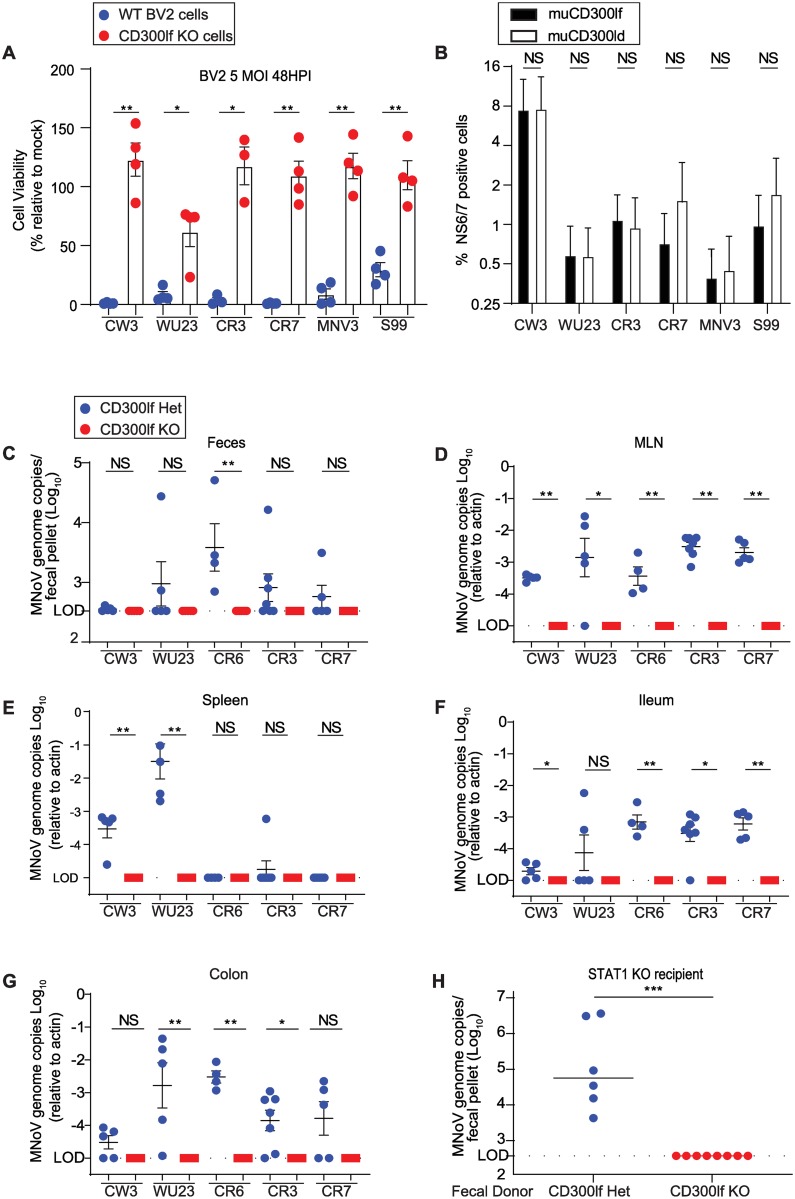
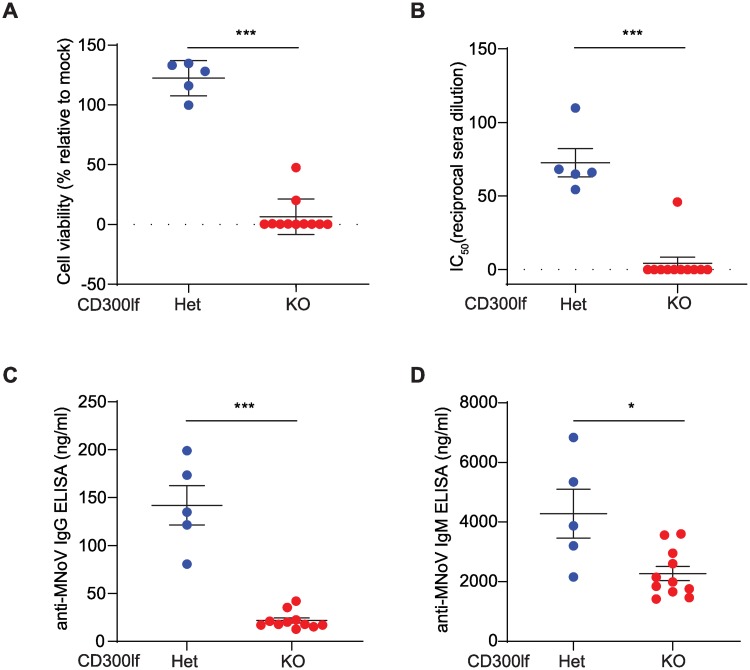
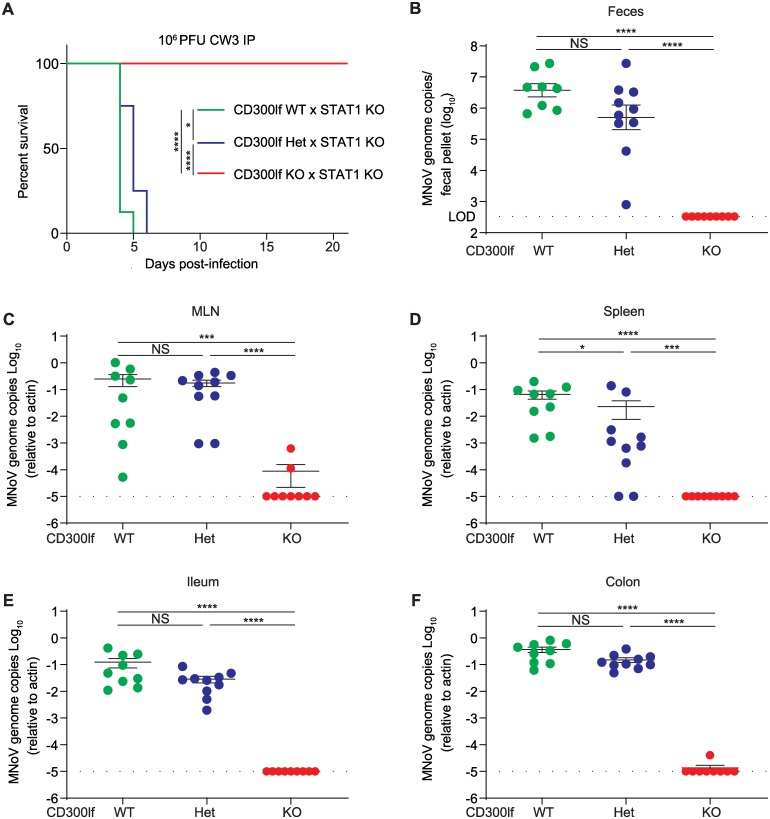
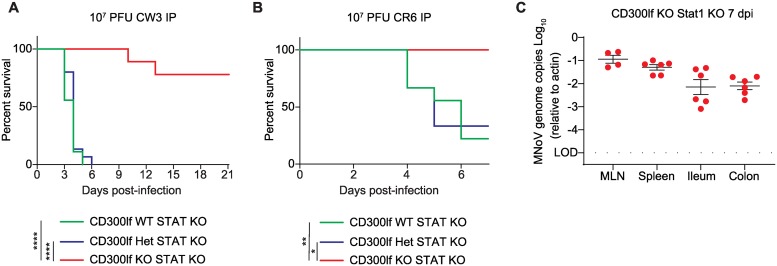
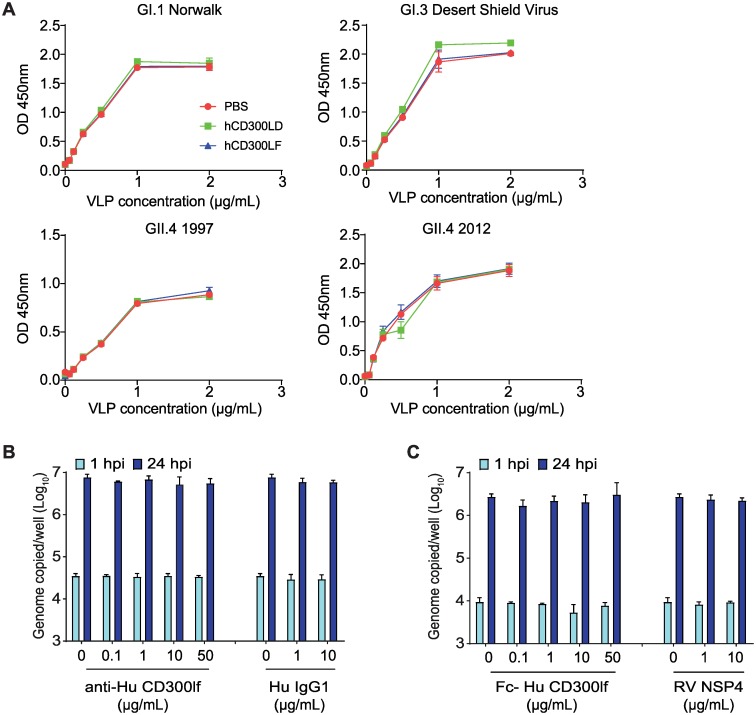
Murine norovirus (MNoV) is an important model of human norovirus (HNoV) and mucosal virus infection more broadly. Viral receptor utilization is a major determinant of cell tropism, host range, and pathogenesis. The bona fide receptor for HNoV is unknown. Recently, we identified CD300lf as a proteinaceous receptor for MNoV. Interestingly, its paralogue CD300ld was also sufficient for MNoV infection in vitro. Here we explored whether CD300lf is the sole physiologic receptor in vivo and whether HNoV can use a CD300 ortholog as an entry receptor. We report that both CD300ld and CD300lf are sufficient for infection by diverse MNoV strains in vitro. We further demonstrate that CD300lf is essential for both oral and parenteral MNoV infection and to elicit anti-MNoV humoral responses in vivo. In mice deficient in STAT1 signaling, CD300lf is required for MNoV-induced lethality. Finally, we demonstrate that human CD300lf (huCD300lf) is not essential for HNoV infection, nor does huCD300lf inhibit binding of HNoV virus-like particles to glycans. Thus, we report huCD300lf is not a receptor for HNoV.
Conflict of interest statement
CBW and RCO are inventors on a patent application submitted by Washington University entitled “Receptor for norovirus and uses thereof” (U.S. Provisional Application 62/301,965).
Figures
Similar articles
-
CD300lf Conditional Knockout Mouse Reveals Strain-Specific Cellular Tropism of Murine Norovirus.J Virol. 2021 Jan 13;95(3):e01652-20. doi: 10.1128/JVI.01652-20. Print 2021 Jan 13. J Virol. 2021. PMID: 33177207 Free PMC article.
-
Functional receptor molecules CD300lf and CD300ld within the CD300 family enable murine noroviruses to infect cells.Proc Natl Acad Sci U S A. 2016 Oct 11;113(41):E6248-E6255. doi: 10.1073/pnas.1605575113. Epub 2016 Sep 28. Proc Natl Acad Sci U S A. 2016. PMID: 27681626 Free PMC article.
-
Atomic Structure of the Murine Norovirus Protruding Domain and Soluble CD300lf Receptor Complex.J Virol. 2018 May 14;92(11):e00413-18. doi: 10.1128/JVI.00413-18. Print 2018 Jun 1. J Virol. 2018. PMID: 29563286 Free PMC article.
-
Norovirus Attachment and Entry.Viruses. 2019 May 30;11(6):495. doi: 10.3390/v11060495. Viruses. 2019. PMID: 31151248 Free PMC article. Review.
-
Norovirus Regulation by Host and Microbe.Trends Mol Med. 2016 Dec;22(12):1047-1059. doi: 10.1016/j.molmed.2016.10.003. Epub 2016 Nov 22. Trends Mol Med. 2016. PMID: 27887808 Free PMC article. Review.
Cited by
-
Tuft Cells: Context- and Tissue-Specific Programming for a Conserved Cell Lineage.Annu Rev Pathol. 2023 Jan 24;18:311-335. doi: 10.1146/annurev-pathol-042320-112212. Epub 2022 Nov 9. Annu Rev Pathol. 2023. PMID: 36351364 Free PMC article. Review.
-
Genetic diversity of murine norovirus populations less susceptible to chlorine.Front Microbiol. 2024 Apr 30;15:1372641. doi: 10.3389/fmicb.2024.1372641. eCollection 2024. Front Microbiol. 2024. PMID: 38746753 Free PMC article.
-
Lipid nanoparticles as adjuvant of norovirus VLP vaccine augment cellular and humoral immune responses in a TLR9- and type I IFN-dependent pathway.J Virol. 2024 Dec 17;98(12):e0169924. doi: 10.1128/jvi.01699-24. Epub 2024 Nov 4. J Virol. 2024. PMID: 39494905 Free PMC article.
-
Selective Polyprotein Processing Determines Norovirus Sensitivity to Trim7.J Virol. 2022 Sep 14;96(17):e0070722. doi: 10.1128/jvi.00707-22. Epub 2022 Aug 16. J Virol. 2022. PMID: 35972292 Free PMC article.
-
Multimodal control of dendritic cell functions by nociceptors.Science. 2023 Mar 31;379(6639):eabm5658. doi: 10.1126/science.abm5658. Epub 2023 Mar 31. Science. 2023. PMID: 36996219 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
