

Outer Membrane Lipid Secretion and the Innate Immune Response to Gram-Negative Bacteria
- PMID: 32253250
- PMCID: PMC7309610
- DOI: 10.1128/IAI.00920-19
Outer Membrane Lipid Secretion and the Innate Immune Response to Gram-Negative Bacteria
Abstract
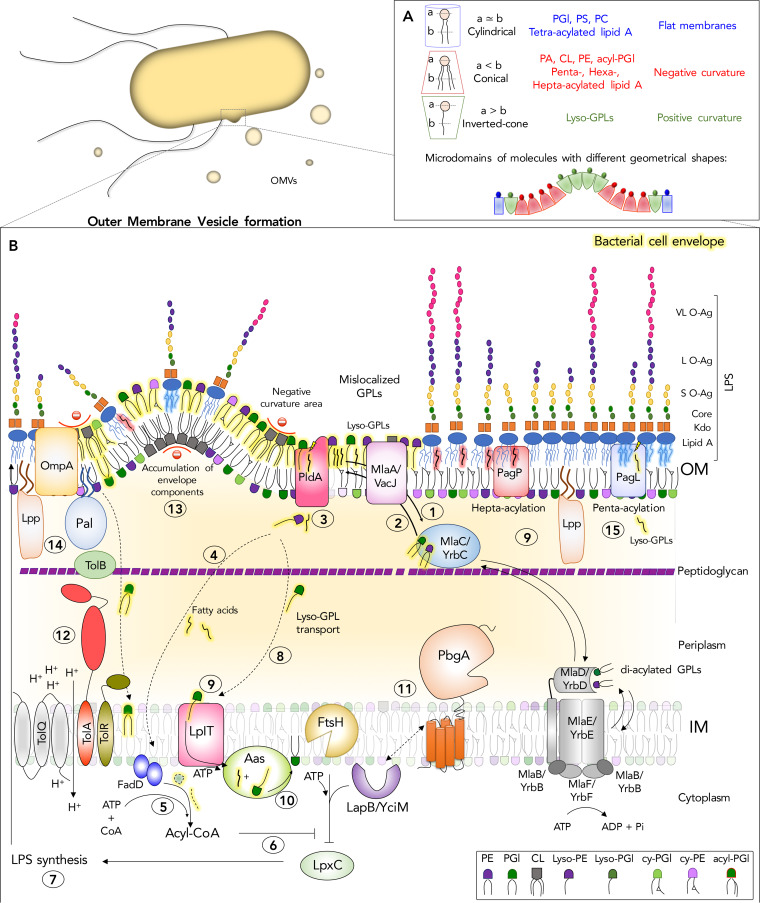
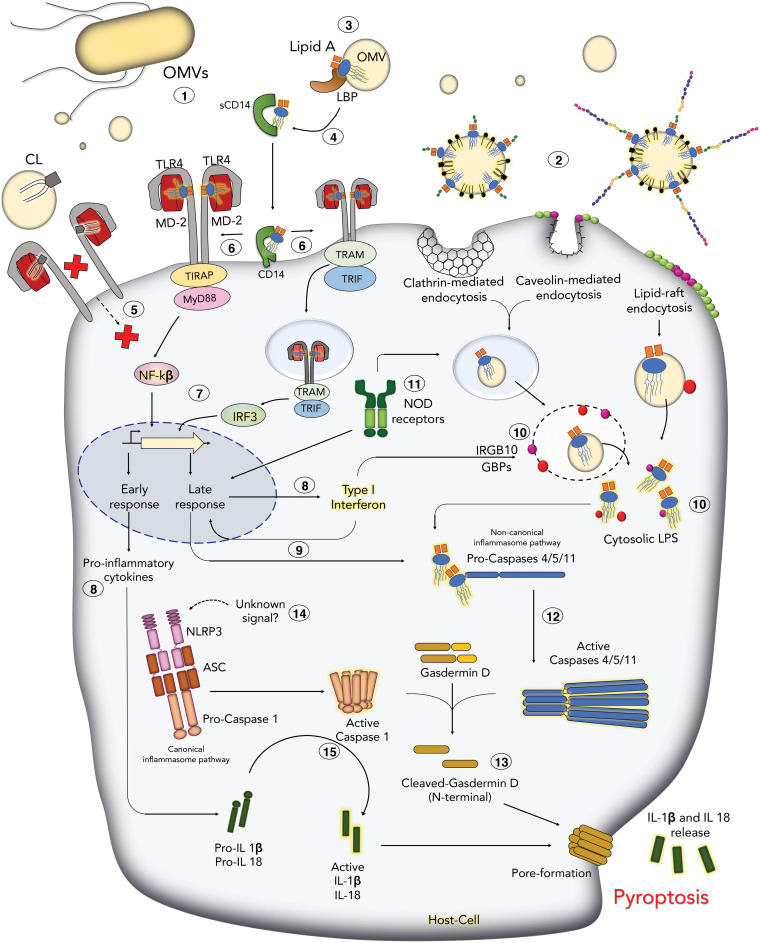
The outer membrane (OM) of Gram-negative bacteria is an asymmetric lipid bilayer that consists of inner leaflet phospholipids and outer leaflet lipopolysaccharides (LPS). The asymmetric character and unique biochemistry of LPS molecules contribute to the OM's ability to function as a molecular permeability barrier that protects the bacterium against hazards in the environment. Assembly and regulation of the OM have been extensively studied for understanding mechanisms of antibiotic resistance and bacterial defense against host immunity; however, there is little knowledge on how Gram-negative bacteria release their OMs into their environment to manipulate their hosts. Discoveries in bacterial lipid trafficking, OM lipid homeostasis, and host recognition of microbial patterns have shed new light on how microbes secrete OM vesicles (OMVs) to influence inflammation, cell death, and disease pathogenesis. Pathogens release OMVs that contain phospholipids, like cardiolipins, and components of LPS molecules, like lipid A endotoxins. These multiacylated lipid amphiphiles are molecular patterns that are differentially detected by host receptors like the Toll-like receptor 4/myeloid differentiation factor 2 complex (TLR4/MD-2), mouse caspase-11, and human caspases 4 and 5. We discuss how lipid ligands on OMVs engage these pattern recognition receptors on the membranes and in the cytosol of mammalian cells. We then detail how bacteria regulate OM lipid asymmetry, negative membrane curvature, and the phospholipid-to-LPS ratio to control OMV formation. The goal is to highlight intersections between OM lipid regulation and host immunity and to provide working models for how bacterial lipids influence vesicle formation.
Keywords: 4; MD-2; OMV; TLR4; Tol-Pal; Toll-like receptor; and 5; antibiotic resistance; antibiotics; antimicrobial peptides; asymmetry; bleb; cardiolipin; caspases 11; constriction; endocytosis; endotoxin; ftsH; gasdermin; glycerophospholipid; immunity; inflammasome; inflammation; lapB/yciM; lipid A; lipid rafts; lipooligosaccharides; lipopolysaccharide; lpp; lpxC; microbial associated molecular patterns; mla; myeloid differentiation factor 2; ompA; outer membrane vesicles; pagL; pagP; pattern recognition receptor; pbgA; peptidoglycan; permeability barrier; pldA; pyroptosis; secretion systems; yejM.
Copyright © 2020 American Society for Microbiology.
Figures
References
-
- Gioannini TL, Teghanemt A, Zhang D, Coussens NP, Dockstader W, Ramaswamy S, Weiss JP. 2004. Isolation of an endotoxin-MD-2 complex that produces Toll-like receptor 4-dependent cell activation at picomolar concentrations. Proc Natl Acad Sci U S A 101:4186–4191. doi: 10.1073/pnas.0306906101. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
