

Studying heat shock proteins through single-molecule mechanical manipulation
- PMID: 32253740
- PMCID: PMC7332600
- DOI: 10.1007/s12192-020-01096-y
Studying heat shock proteins through single-molecule mechanical manipulation
Abstract
Imbalances of cellular proteostasis are linked to ageing and human diseases, including neurodegenerative and neuromuscular diseases. Heat shock proteins (HSPs) and small heat shock proteins (sHSPs) together form a crucial core of the molecular chaperone family that plays a vital role in maintaining cellular proteostasis by shielding client proteins against aggregation and misfolding. sHSPs are thought to act as the first line of defence against protein unfolding/misfolding and have been suggested to act as "sponges" that rapidly sequester these aberrant species for further processing, refolding, or degradation, with the assistance of the HSP70 chaperone system. Understanding how these chaperones work at the molecular level will offer unprecedented insights for their manipulation as therapeutic avenues for the treatment of ageing and human disease. The evolution in single-molecule force spectroscopy techniques, such as optical tweezers (OT) and atomic force microscopy (AFM), over the last few decades have made it possible to explore at the single-molecule level the structural dynamics of HSPs and sHSPs and to examine the key molecular mechanisms underlying their chaperone activities. In this paper, we describe the working principles of OT and AFM and the experimental strategies used to employ these techniques to study molecular chaperones. We then describe the results of some of the most relevant single-molecule manipulation studies on HSPs and sHSPs and discuss how these findings suggest a more complex physiological role for these chaperones than previously assumed.
Keywords: Heat shock proteins; Mechanism of action; Single-molecule manipulation; Small heat shock proteins; Structural dynamics.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
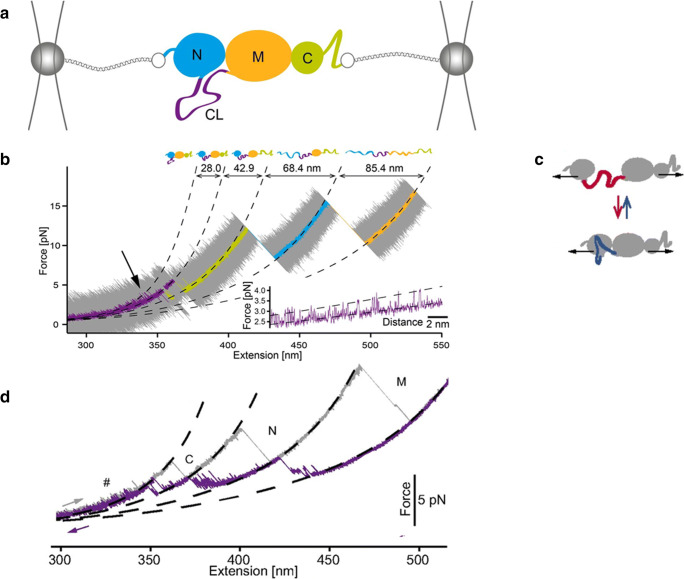
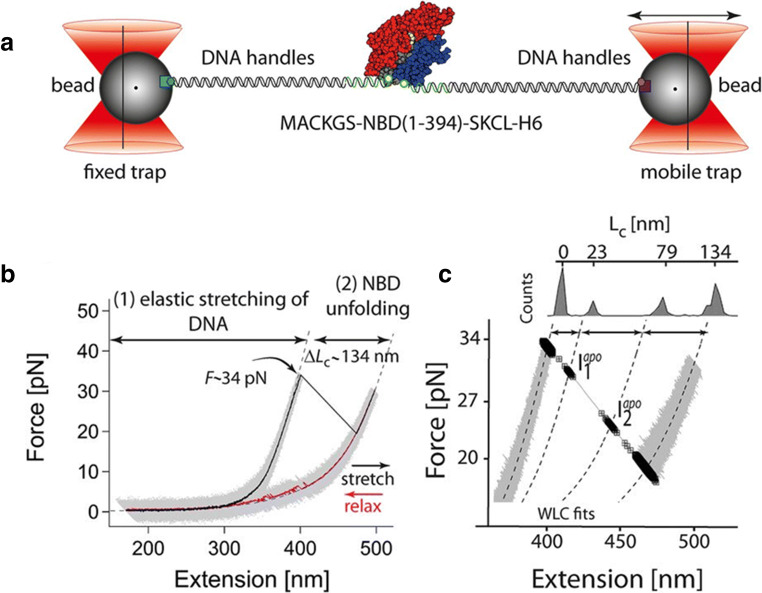
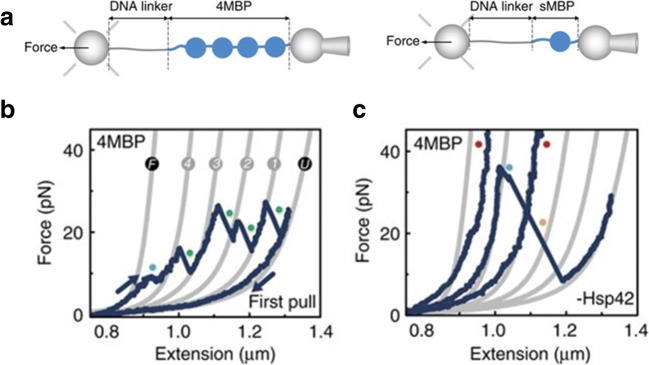
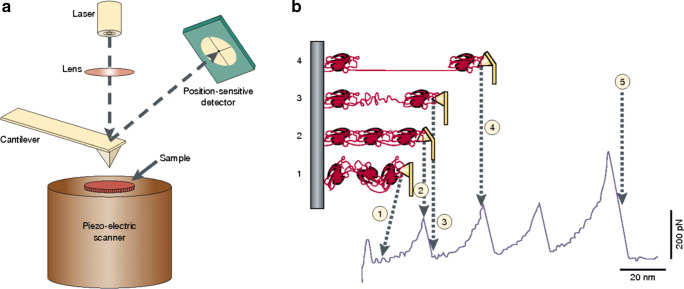
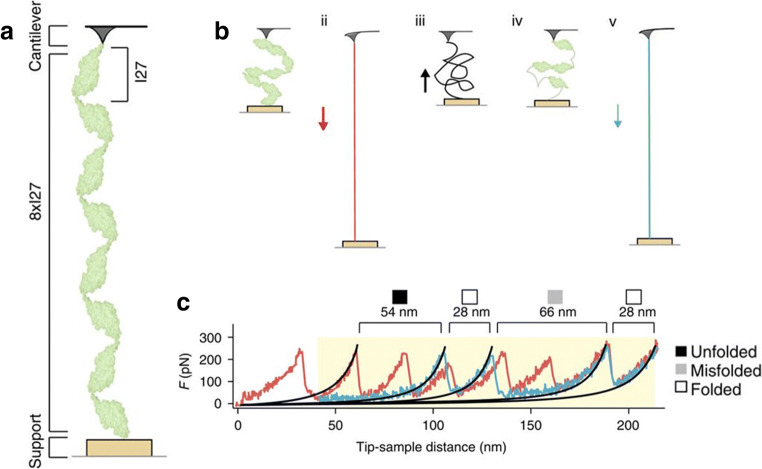
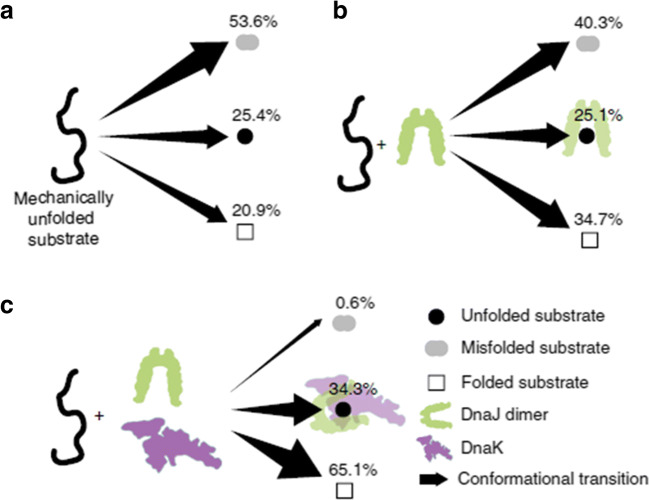
References
-
- Balchin D, Hayer-Hartl M, Hartl FU. In vivo aspects of protein folding and quality control. Science. 2016;353(6294):aac4354. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
